Skip to comments.
IMAX steers clear of Darwin's theory
WorldNetDaily ^
| 3/20/05
| WorldNetDaily
Posted on 03/20/2005 12:01:05 PM PST by wagglebee
Some IMAX theaters are refusing to carry movies that promote evolution, citing concerns that doing so offends their audience and creates controversy – a move that has some proponents of Darwinism alarmed over the influence of "fundamentalists."
It's a decision that affects not only the network of 240 IMAX theaters operating in 35 countries, but some science museums that show IMAX-formatted films.
IMAX, which bills itself as the "ultimate movie experience," promises to take viewers to "places you only imagined." The 8-story high screens and crystal clear images have made the theaters ideal venues for documentary science films showing the splendor of nature.
Now, however, about a dozen IMAX theaters, primarily in the South, are shunning movies that carry evolution themes, the New York Times reports. Fear of protests by those objecting to films that contradict the Biblical account of creation is cited as the reason.
A dozen science centers rejected the 2003 release, "Volcanoes," because of it speculation that life on Earth may have originated in undersea vents, says Dr. Richard Lusk, an oceanographer and chief scientist for the project.
Because a only small number of IMAX theaters show science films, a boycott by a few can reduce the potential audience to the point that producers question whether projects are financially worthwhile.
"We have definitely a lot more creation public than evolution public," says Lisa Buzzelli, of the Charleston, South Carolina, Imax Theater. "Being in the Bible Belt, ["Volcanoes"] does have a lot to do with evolution, and we weigh that carefully."
When the Fort Worth Museum of Science and History played the movie for a test audience, the responses were sufficiently negative for the museum to drop it from its offerings. Responses like "I really hate it when the theory of evolution is presented as fact," or "I don't agree with their presentation of human existence" doomed the film's chances.
"Some people said it was blasphemous," says Carol Murray, the museum's director of marketing. "If it's not going to draw a crowd and it is going to create controversy," she concludes, "from a marketing standpoint I cannot make a recommendation" to screen it.
The film's distributor says other science museum officials turned him down "for religious reasons" and because "Volcanoes" had "evolutionary overtones" – a claim that makes Hyman Field, a former National Science Foundation official who played a role in its financing, "furious."
"It's very alarming," he says, "all of this pressure being put on a lot of the public institutions by the fundamentalists."
The economics of large-format science documentaries being what they are, it might not take too much pressure for filmmakers to begin avoiding Darwin.
The films "are generally not big moneymakers," notes Joe DeAmicis, former director of the IMAX theater at the California Science Center in Los Angeles. "It's going to be hard for our filmmakers to continue to make unfettered documentaries when they know going in that 10 percent of the market" will reject them.
Bayley Silleck, who wrote and directed "Cosmic Voyage," another IMAX offering that drew religious complaints, expects to encounter criticism on his upcoming project about dinosaurs. While he's critical of "overcaution, overprotectedness" by theater operators, he recognizes that in the end, it's the audience that counts.
"We all have to make films for an audience that is a family audience," he observes, "when you are talking about IMAX, because they are in science centers and museums."
A Gallup poll, released earlier this month, reveals that 81 percent of U.S. teenagers believe God was somehow involved in human origins, with only 18 percent holding a purely secular view of evolution.
TOPICS:
KEYWORDS: bible; creationism; crevolist; darwin; evolution; gallup; imax; movies; religion; science; secularhumanism; secularism
Navigation: use the links below to view more comments.
first previous 1-20 ... 121-140, 141-160, 161-180 ... 221-236 next last
To: Terriergal
How about that Anthony Flew quotation, with source? I believe this is the third time I'm requesting it.
You know, it looks bad to simply make up stuff like that.
Some would even call it a sin.
To: Tamberlane
See my post #135.
It just seems to me that scientists who believe in the theory of evolution simply change the story when new findings disprove what they have previously believed.
To: Prov3456
Your post #135 is what I was replying to.
To: Oztrich Boy
OK, but that's YOUR problem, not Ken Ham's.
Just because you do not like him, or his positions, does not mean that they are false (or true).
To: Prov3456
"It just seems to me that scientists who believe in the theory of evolution simply change the story when new findings disprove what they have previously believed."
Thats what science does. You try to come up with the best explanation you can with the evidence you have. If the evidence changes, you change your explanation so that the new data fits. Thats why It's so funny when creationists say that science ignores "facts". Science is all about "accepting new data".
As opposed to creationists who start "knowing the truth" and try to find evidence to back it up.
145
posted on
03/20/2005 2:27:46 PM PST
by
ndt
To: Prov3456
[Now perhaps you'd care to retract your unfounded slur on me.] No, I'm not ready to do that yet.
Color me unsurprised.
But I will offer some reading materials for your consideration:
Icons of Evolution: Science or Myth by Jonathan Wells
Read it already. Full of misrepresentations and misunderstandings on Wells's part. Classic "straw man fallacy" stuff (i.e. beating up a sham scarecrow replica of your opponent's position and then declaring "victory" over his *actual* position), as well as countless outright falsehoods. I haven't bothered to write my own review of it because this webpage already does such a good job of expressing my own opinion of the book: Icons of Evolution FAQs, especially in (but not limited to) this sub-page: Icon of Obfuscation Jonathan Wells's book Icons of Evolution: and why most of what it teaches about evolution is wrong. I don't just take Matzke's word for it -- I can personally vouch for the accuracy of his refutations of Wells's flawed points.
Of Pandas and People: The Central Question of Biological Origins by Dean H. Kenyon (biologist) and Percival Davis (zoologist). It compares the theories of evolution and "intelligent design" but does not mention God, Christ, the Bible, church or creation. It is a textbook that was reviewed by 35 reviewers, including evolutionists and non-evolutionists.
Haven't read it. But the full text is searchable and readable on Amazon.com, and when I tried a couple of keywords just now ("DNA" and "Cambrian"), I found a lot of the usual creationist misrepresentations and misunderstandings, so I can't say that I'm impressed. For example, he's just monumentally wrong (and jaw-droppingly ignorant) when he claims that:
This nearly simultaneous appearance of most known phyla [during the Cambrian - Ich.] is more remarkable when we consider that the variation within a phylum is quite small compared to how much the phyla vary from one another. In other words, there is more morphological distance between two phyla than separates representatives within the phyla themselves. This means that the origins of new phyla are evolution's greatest achievements in diversifying life forms."
Um, no. The author is making the ludicrous claim that there is *LESS* structural/evolutionary difference between, say, a parrot and a hagfish:

 ...(both are members of the chordata phylum) than between a Cambrian worm with a primitive notocord versus a Cambrian worm with a more diffuse neural net. Nice try.
...(both are members of the chordata phylum) than between a Cambrian worm with a primitive notocord versus a Cambrian worm with a more diffuse neural net. Nice try.
The DNA material was equally giggle-worthy.
If you can direct me to a page number you feel makes a decent point without such serious flaws, let me know and I'll check it out.
Darwin on Trial by Phillip Johnson
Defeating Darwinism by Opening Minds by Phillip Johnson
I've read the former, and based on its worthlessness I skipped reading the latter. I also debated Johnson online back and forth for a week about ten years ago. What he doesn't know about biology would fill volumes. He arguest against evolution in exactly the way you would expect him to as a lawyer (his actual profession) -- by using what *sounds* persuasive instead of on what is actually sound reasoning, or actually founded upon the preponderance of the evidence. I wasn't at all impressed. And apparently I'm not the only one. See for example:
The Truth, the Whole Truth, and Nothing but the Truth? Why Phillip Johnson's Darwin on Trial and the "Intelligent Design" movement are neither science—nor Christian
Critiques of Anti-Evolutionist Phillip Johnson's Views
DARWIN ON TRIAL: A Review
Darwin's Black Box by Michael Behe (biologist)
I've read that too. Behe seems sincere enough, at least, but in his zeal he produces shoddy, flawed work, while wildly overstating what he can actually support (if at all). Here are some of my prior posts on the problems in Behe's book and other statements/publications:
The next idea you probably will not like, and that is irreducible complexity.
As an "idea" I like it just fine, and so do evolutionary scientists. The problem is that Behe (and the creationists who follow him) have created a "straw man" version of "IC" which is quite simply incorrect -- but appears to give the conclusion they want.
The original notion of "IC" goes back to Darwin himself. He wrote:
"If it could be demonstrated that any complex organ existed, which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down."
-- Charles Darwin, "On the Origin of Species", 1859
That's "Irreducible Complexity" in a nutshell. It's not as if Behe has pointed out anything that biologists (or Darwin) didn't already realize. But let's examine Darwin's description of "IC" in a bit more detail (emphasis mine):
No doubt many organs exist of which we do not know the transitional grades, more especially if we look to much-isolated species, round which, according to my theory, there has been much extinction. Or again, if we look to an organ common to all the members of a large class, for in this latter case the organ must have been first formed at an extremely remote period, since which all the many members of the class have been developed; and in order to discover the early transitional grades through which the organ has passed, we should have to look to very ancient ancestral forms, long since become extinct. We should be extremely cautious in concluding that an organ could not have been formed by transitional gradations of some kind. Numerous cases could be given amongst the lower animals of the same organ performing at the same time wholly distinct functions; thus the alimentary canal respires, digests, and excretes in the larva of the dragon-fly and in the fish Cobites. In the Hydra, the animal may be turned inside out, and the exterior surface will then digest and the stomach respire. In such cases natural selection might easily specialise, if any advantage were thus gained, a part or organ, which had performed two functions, for one function alone, and thus wholly change its nature by insensible steps. Two distinct organs sometimes perform simultaneously the same function in the same individual; to give one instance, there are fish with gills or branchiae that breathe the air dissolved in the water, at the same time that they breathe free air in their swimbladders, this latter organ having a ductus pneumaticus for its supply, and being divided by highly vascular partitions. In these cases, one of the two organs might with ease be modified and perfected so as to perform all the work by itself, being aided during the process of modification by the other organ; and then this other organ might be modified for some other and quite distinct purpose, or be quite obliterated.
The illustration of the swimbladder in fishes is a good one, because it shows us clearly the highly important fact that an organ originally constructed for one purpose, namely flotation, may be converted into one for a wholly different purpose, namely respiration. The swimbladder has, also, been worked in as an accessory to the auditory organs of certain fish, or, for I do not know which view is now generally held, a part of the auditory apparatus has been worked in as a complement to the swimbladder. All physiologists admit that the swimbladder is homologous, or 'ideally similar,' in position and structure with the lungs of the higher vertebrate animals: hence there seems to me to be no great difficulty in believing that natural selection has actually converted a swimbladder into a lung, or organ used exclusively for respiration.
[Example snipped]
In considering transitions of organs, it is so important to bear in mind the probability of conversion from one function to another, that I will give one more instance. [Long detail of example snipped] If all pedunculated cirripedes had become extinct, and they have already suffered far more extinction than have sessile cirripedes, who would ever have imagined that the branchiae in this latter family had originally existed as organs for preventing the ova from being washed out of the sack?
-- Charles Darwin, "On the Origin of Species", 1859
Darwin makes two critical points here: 1. A modern organ need not have evolved into its present form and function from a precursor which had always performed the same function. Evolution is quite capable of evolving a structure to perform one function, and then turning it to some other "purpose".
2. Organs/structures can reach their present form through a *loss* of function or parts, not just through *addition* of function or parts.
Despite the fact that these observations were laid out in 1859, Behe's version of "Irreducible Complexity" pretends they are not factors, and defines "IC" as something which could not have arisen through stepwise *ADDITIONS* (only) while performing the same function *THROUGHOUT ITS EXISTENCE*.
It's hard to tell whether Behe does this through ignorance or willful dishonesty, but the fact remains that *his* definition and analysis of "IC" is too restrictive. He places too many "rules" on how he will "allow" evolution to reach his examples of "Behe-style IC" structures, while evolution itself *IS NOT RESTRICTED TO THOSE RULES* when it operates. Thus Behe's conclusion that "Behe-style evolution" can not reach "Behe-style IC" hardly tells us anything about whether *real-world* evolution could or could not have produced them.
For specific examples, Behe's example of the "Behe-style IC" flagellum is flawed because flagella are composed of components that bacteria use FOR OTHER PURPOSES and were evolved for those purposes then co-opted (1, 2), and Behe's example of the "Behe-style IC" blood-clotting process is flawed because the biochemistry of blood-clotting is easily reached by adding several steps on top of a more primitive biochemical sequence, *and then REMOVING earlier portions which had become redundant* (1, 2).
Even Behe's trivial mousetrap example turns out to not actually be "IC".
The usual qualitative formulation is: "An irreducibly complex system cannot be produced...by slight, successive modifications of a precursor system, because any precursor to an irreducibly complex system, that is missing a part is by definition nonfunctional..."
Note the key error: By saying that it "breaks" if any part is "missing" (i.e. taken away), it is only saying that evolution could not have reached that endpoint by successively only ADDING parts. True enough, but Behe misses the fact that you can also reach the same state by, say, adding 5 parts one at a time, and then taking away 2 which have become redundant. Let's say that part "A" does the job, but not well. But starting with just "A" serves the need. Then add "B", which improves the function of "A". Add "C" which helps A+B do their job, and so on until you have ABCDE, which does the job very well. Now, however, it may turn out that CDE alone does just fine (conceivably, even better than ABCDE does with A+B getting in the way of CDE's operation). So A and B fade away, leaving CDE. Note that CDE was built in "one change at a time" fashion, with each new change improving the operation. HOWEVER, by Behe's definition CDE is "Irreducibly Complex" and "could not have evolved (been built by single steps)" because removing C or D or E from CDE will "break" it. Note that Behe's conclusion is wrong. His logic is faulty.
The other error in Behe's definition lies in this part: "...any precursor to an irreducibly complex system, that is missing a part is by definition nonfunctional". The problem here is that it may be "nonfunctional" for its *current* function, but perfectly functional for some *other* function helpful for survival (and therefore selected by evolution). Behe implicitly claims that if it's not useful for its *current* function, it's useless for *any* function. The flaw in this should be obvious.
"Since natural selection can only choose systems that are already working, then if a biological system cannot be produced gradually it would have to arise as an integrated unit, in one fell swoop, for natural selection to have anything to act on."
True as far as it goes, but but this is hardly the same as Behe's sleight-of-hand in the first part of his statement, which relies on the false premise that a precursor to a structure is 100% useless for *any* purpose if *taking away* (but not adding) one part from the current purpose makes it unsuitable for the current purpose. Two gaping holes in that one...
Behe (an anathematized name)
For reasons I've outlined above.
talks of the bacterial flagellum, which contains an acid-powered rotary engine, a stator, O-rings, bushings, and a drive shaft. The machinery of this motor requires approximately fifty proteins.
Except that it doesn't. As many biochemists have pointed out, other organisms have function flagella (even *as* flagella) with fewer proteins (and/or different proteins). That flagellum isn't even "IC" by Behe's own definition since you *can* remove proteins and have it still work as a flagellum. [...]
For a far more realistic look at the evolutionary "invention" of the flagellum, see Evolution in (Brownian) space: a model for the origin of the bacterial flagellum , which I linked earlier in this post. From the abstract:
The model consists of six major stages: export apparatus, secretion system, adhesion system, pilus, undirected motility, and taxis-enabled motility. The selectability of each stage is documented using analogies with present-day systems. Conclusions include: (1) There is a strong possibility, previously unrecognized, of further homologies between the type III export apparatus and F1F0-ATP synthetase. (2) Much of the flagellum’s complexity evolved after crude motility was in place, via internal gene duplications and subfunctionalization. (3) Only one major system-level change of function, and four minor shifts of function, need be invoked to explain the origin of the flagellum; this involves five subsystem-level cooption events. (4) The transition between each stage is bridgeable by the evolution of a single new binding site, coupling two pre-existing subsystems, followed by coevolutionary optimization of components. Therefore, like the eye contemplated by Darwin, careful analysis shows that there are no major obstacles to gradual evolution of the flagellum.
And:
For an analysis of numerous errors and such in Dembski's Design arguments/examples, see Not a Free Lunch But a Box of Chocolates: A critique of William Dembski's book No Free Lunch. It also contains material on the flagella issue you raise next.
As for Behe (the other author):
One small example is the flagella on a paramecium. They need four distinct proteins to work.
Actually they need a lot more than that. And as far as I know, Behe never used the cilia on paramecia as his example, he has primarily concentrated on bacterial flagella.
They cannot have evolved from a flagella that need three.
Contrary to creationist claims (or Behe's) that flagella are Irreducibly Complex and can not function at all if any part or protein is removed, in fact a) there are many, many varieties of flagella on various species of single-celled organisms, some with more or fewer parts/proteins than others. So it's clearly inaccurate to make a blanket claim that "flagella" in general contain no irreplacable parts. Even Behe admits that a working flagella can be reduced to a working cilia, which undercuts his entire "Irreducibly Complex" example/claim right off the bat.
For a semi-technical discussion of how flagella are *not* IC, because many of their parts can be eliminated without totally breaking their locomotive ability, see Evolution of the Bacterial Flagella
But even if one could identify, say, four specific proteins (or other components) which were critically necessary for the functioning of all flagellar structures (and good luck: there are three unrelated classes of organisms with flagella built on three independent methods: eubacterial flagella, archebacterial flagella, and eukaryote flagella -- see Faugy DM and Farrel K, (1999 Feb) A twisted tale: the origin and evolution of motility and chemotaxis in prokaryotes. Microbiology, 145, 279-280), Behe makes a fatal (and laughably elementary) error when he states that therefore they could not have arisen by evolution. Even first-year students of evolutionary biology know that quite often evolved structures are built from parts that WERE NOT ORIGINALLY EVOLVED FOR THEIR CURRENT APPLICATION, as Behe naively assumes (or tries to imply).
Okay, fine, so even if you can prove that a flagellum needs 4 certain proteins to function, and would not function AS A FLAGELLUM with only 3, that's absolutely no problem for evolutionary biology, since it may well have evolved from *something else* which used those 3 proteins to successfully function, and only became useful as a method of locomotion when evolution chanced upon the addition of the 4th protein. Biology is chock-full of systems cobbled together from combinations of other components, or made via one addition to an existing system which then fortuitously allows it to perform a new function.
And, lo and behold, it turns out that the "base and pivot" of the bacterial flagella, along with part of the "stalk", is virtually identical to the bacterial Type III Secretory Structure (TTSS). So despite Behe's claim that flagella must be IC because (he says) there's no use for half a flagella, in fact there is indeed such a use. And this utterly devastates Behe's argument, in several different ways. Explaining way in detail would take quite some time, but it turns out that someone has already written an excellent essay on that exact thing, which I strongly encourage you to read: The Flagellum Unspun: The Collapse of "Irreducible Complexity" .
(Note: Several times that essay makes a reference to the "argument from ignorance", with the assumption that the reader is already familiar with it. I'd like to point out that contrary to the way it sounds, Miller is *not* accusing Behe et all of being ignorant. Instead, he's referring to this family of logical fallacies, also known as the "argument from incredulity".)
That is called irreducible complexity.
That's what Behe likes to call it, yes. But the flagella is provably *not* IC. Oops for Behe. Furthermore, while it's certainly easy to *call* something or another "Irreducibly Complex", proving that it actually *is* is another matter entirely.
As the "Flagellum Unspun" article above states:
According to Dembski, the detection of "design" requires that an object display complexity that could not be produced by what he calls "natural causes." In order to do that, one must first examine all of the possibilities by which an object, like the flagellum, might have been generated naturally. Dembski and Behe, of course, come to the conclusion that there are no such natural causes. But how did they determine that? What is the scientific method used to support such a conclusion? Could it be that their assertions of the lack of natural causes simply amount to an unsupported personal belief? Suppose that there are such causes, but they simply happened not to think of them? Dembski actually seems to realize that this is a serious problem. He writes: "Now it can happen that we may not know enough to determine all the relevant chance hypotheses [which here, as noted above, means all relevant natural processes (hvt)]. Alternatively, we might think we know the relevant chance hypotheses, but later discover that we missed a crucial one. In the one case a design inference could not even get going; in the other, it would be mistaken" (Dembski 2002, 123 (note 80)).
For more bodyblows against the notion of Irreducible Complexity, see: Bacterial Flagella and Irreducible Complexity
Irreducible Complexity Demystified
Irreducible Complexity
Review: Michael Behe's "Darwin's Black Box"
The fatal flaws in Behe's argument were recognized as soon as his book was published, and countless reviewers pointed them out. And yet, creationists and IDers, who seem to rely mostly on the echo-chamber of their own clique and appear to seldom read much *actual* scientific sources, still seem blissfully unaware of the problems with Behe's thesis, and keep popping in on a regular basis to wave the book around and smugly yell something like, "See, evolution has already been disproven!"
What's funny is that by Behe's own argument, a stone arch is "irreducibly complex" because it could not have formed by nature *adding* sections of stone at a time (it would have fallen down unless the entire span was already in place -- and indeed will fall down if you take part of the span away):

Needless to say, what Behe's argument is missing in the case of the stone arch is that such arches form easily by natural means when successive layers of sedimentary rock added on top of each other, and *then* erosion carves a hole out from *under* the arch by *removing* material after the "bridge" of the arch itself *was already there*.
Similarly, Behe's arguments about why certain types of biological structures "could not" have evolved fall flat because he doesn't realize that evolution does not only craft features by *adding* components, it also does so by *lateral alteration*, and by *removing* components.
Behe's "irreducible complexity" argument is fatally flawed. It only "proves" that a *simplified* version of evolution (as envisioned by Behe) couldn't give rise to certain structures -- not that the *actual* processes of evolution could not.
Evolution: A Theory in Crisis by Michael Denton (biologist).
Already read it. More misrepresentations and misunderstandings about what evolutionary biology *actually* consists of and the nature of the evidence supporting it. One example from a prior post of mine:
Michael Denton, an Australian biologist and self-described agnostic. Denton writes that evolutionists once thought that comparing DNA sequences would prove the "family tree" linkage between species that Darwin conceived. But "Thousands of different sequences, protein and nucleic acid, have now been compared in hundreds of different species, but never has any sequence been found to be in any sense the lineal descendant or ancestor of any other sequence,"
To be blunt, Denton is either an idiot or a liar. His claim is flat wrong. For many specific examples of five entirely *independent* methods of linking common ancestry via DNA analysis, see Molecular Sequence Evidence. For *tons* of research studies turning up more DNA evidence of common ancestry on a regular basis, see The Journal of Molecular Biology. You can browse abstracts from hundreds of articles publshed in the past 89 issues on that site. For full text, subscribe to the online version or go visit a technical library. From just the most recent issue, for example:
The PRAT Purine Synthesis Gene Duplication in Drosophila melanogaster and Drosophila virilis Is Associated with a Retrotransposition Event and Diversification of Expression Patterns (short summary: the authors identified gene sequences which were inherited from a common ancestor of the two species 40 million years ago) Phylogeny of Choanozoa, Apusozoa, and Other Protozoa and Early Eukaryote Megaevolution (short summary: A study of DNA sequences and the light it sheds on the very early split of the various single-cell organism types from a common ancestor)
Frequent Mitochondrial Gene Rearrangements at the Hymenopteran nad3–nad5 Junction (short summary: DNA from 21 distinct groups of wasps were compared and the implications for the family tree and "history" are discussed)
And here's one more from the Journal of Human Genetics: Molecular phylogenetics of the hominoid Y chromosome (short summary: Y-chromosome DNA from humans, chimpanzees, gorillas, and orangutans were compared and the results were as expected if the species share a common ancestor.) Denton is quite simply flat wrong.
Denton also writes, "The complexity of the simplest known type of cell is so great that it is impossible to accept that such an object could have been thrown together by some kind of freakish, vastly improbable event."
Denton's either incompetent or dishonest here, since no one's proposing that any "known type of cell" was representative of the first spark(s) of life. The earliest life was far, far simpler than that. See for example: On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells or The Path from the RNA World
For more negative critiques of Denton's book identifying the errors in his arguments (with which I heartily concur and for which I can vounch), see for example:
Review of Michael Denton's Evolution: A Theory in Crisis
Reviews: "Evolution: A Theory In Crisis" by Michael Denton
I trust this is enough material to begin your search.
Way ahead of you, actually. You might want to read the above material and links in order to catch up with me, however.
Have you got any material that *isn't* obviously seriously flawed? I'll be happy to check it out.
Good reading. (And I mean that sincerely, not sarcastically!)
I wish I could say that they *had* been "good reading", but unfortunately I don't enjoy reading flawed material and misrepresentations of a field about which there is already far too much misinformation already.
I've read a *ton* of creationist (and generically "anti-evolution") literature. But how many primary sources in the scientific literature have you read in comparison to your creationist bookshelf?
To: Ichneumon
Formidable post - my hat is off to you. Thanks for taking the time!
To: Tamberlane
Correct me if I'm wrong, but didn't Darwin's
Origin of the Species postulate that through natural selection and mutation new species would be produced/created?
That, IMO, is the problem. If you'd argue that species change/mutate within themselves, then I'd agree. But it's the idea that monkeys BECAME people (ie. one kind of DNA turning into another kind of DNA) that disproves Darwin's theory for me (and, I suspect, more and more scientists as more is learned about biology, DNA, etc).
Also, I can't quote the source, but I remember reading some time in the past year or so that Darwin returned to his "Christian" roots and rejected his earlier positions on his deathbed. (More food-for-thought-and-conversation!)
To: Terriergal
OH boy. Checking out those links. Yep. Just as I suspected. Only because you "see" what you *want* to see, instead of what's actually there.
It's stuff like this:
[Hovind:] The amount of Helium 4 in the atmosphere, divided by the formation rate on earth, gives only 175,000 years. (God may have created the earth with some helium which would reduce the age more.)
[response:] Notice that the beginnings of this "proof" doesn't support Hovind's claimed Earth age. Hovind then invokes the supernatural to overcome this flaw. LOL It doesn't support Hovind's claimed earth age?
That's correct, it doesn't. Hovind wants to try to "support" an age of the Earth of 6000 years, but his *own argument* produces an age thirty times too large!
Does it support evolution's claimed earth age? 175,000 is a lot closer to 6000 than 6,000,000,000.
Excuse me for being blunt, but if you're not going to even bother to READ the material, why are you attempting to critique it (and thus making a fool of yourself)?
The material you quote out of context GOES ON TO EXPLAIN how and why Hovind's "method" is useless for ANY sort of age determination, and why Hovind's stupid "analysis" reveals a fundamental ignorance on Hovind's part of several of the most BASIC things in the field he's attempting to "analyze".
Try reading for *comprehension* and *learning*, Terriergal, and not for the first *excuse* you can find to hand-wave away the material without risk to your preconceptions.
There is nothing there that actually supports evolution in these links.
Nor did I say that it did. Duh. Hint for the reading-impaired: The links document ERRORS AND DISHONESTIES IN HOVIND'S MATERIAL. It doesn't need to "actually support evolution" in order to identify Hovind's incompetence and dishonesty.
and they will crucify him just as badly as they try to crucify Hovind.
Yawn. No one is "trying to crucify Hovind". We're pointing out the unreliability (to put it mildly) of Hovind's "educational materials".
If you still want to teach your children these known falsehoods, however, that's your choice, and so is the damage you're doing to their education. And to be frank, it could damage their relationship with you and with their faith as well, because eventually they will go out into the world, learn some real science, and discover Mom was feeding them lies about science in the name of God. How do you think they'll react to that?
To: Prov3456
OK, but that's YOUR problem, not Ken Ham's. Just because you do not like him, or his positions, does not mean that they are false (or true). No, of course not. What *does* mean that Hovind's materials are false is the documentation I've linked on this thread.
To: Prov3456
Also, I can't quote the source, but I remember reading some time in the past year or so that Darwin returned to his "Christian" roots and rejected his earlier positions on his deathbed. (More food-for-thought-and-conversation!) I can quote the source, and I remember reading some time in the past year or so that this is one of the fables that Creationists tell themselves to make that Bad Old Science go away.
151
posted on
03/20/2005 3:00:55 PM PST
by
Oztrich Boy
(Rules are for the guidence of wise men, and the blind obedience of fools.)
To: Prov3456
Correct me if I'm wrong, but didn't Darwin's Origin of the Species postulate that through natural selection and mutation new species would be produced/created? Yes he did. And 150 years of subsequent research and evidence has spectacularly confirmed his hypothesis.
That, IMO, is the problem. If you'd argue that species change/mutate within themselves, then I'd agree.
Good.
But it's the idea that monkeys BECAME people (ie. one kind of DNA turning into another kind of DNA) that disproves Darwin's theory for me
Um, why exactly?
(and, I suspect, more and more scientists as more is learned about biology, DNA, etc).
ROFL!! I regret to have to inform you that contrary to your presumptions, the exact opposite is true -- DNA studies and other lines of evidence have overwhelmingly confirmed common descent. For just a small sampling of the evidence for man/ape common ancestry (and there's a *mountain* more like it, and *vast* amounts more concerning the common descent of non-primate life forms), see these previous posts of mine:
Background: Retroviruses reproduce by entering a cell of a host (like, say, a human), then embedding their own viral DNA into the cell's own DNA, which has the effect of adding a "recipe" for manufacturing more viruses to the cell's "instruction book". The cell then follows those instructions because it has no reason (or way) to "mistrust" the DNA instructions it contains. So the virus has converted the cell into a virus factory, and the new viruses leave the cell, and go find more cells to infect, etc.

However, every once in a while a virus's invasion plans don't function exactly as they should, and the virus's DNA (or portions of it) gets embedded into the cell's DNA in a "broken" manner. It's stuck into there, becoming part of the cell's DNA, but it's unable to produce new viruses. So there it remains, causing no harm. If this happens in a regular body cell, it just remains there for life as a "fossil" of the past infection and goes to the grave with the individual it's stuck in. All of us almost certainly contain countless such relics of the past viral infections we've fought off.
However... By chance this sometimes happens to a special cell in the body, a gametocyte cell that's one of the ones responsible for making sperm in males and egg cells in females, and if so subsequent sperm/eggs produced by that cell will contain copies of the "fossil" virus, since now it's just a portion of the entire DNA package of the cell. And once in a blue moon such a sperm or egg is lucky enough to be one of the few which participate in fertilization and are used to produce a child -- who will now inherit copies of the "fossilized" viral DNA in every cell of his/her body, since all are copied from the DNA of the original modified sperm/egg.
So now the "fossilized" viral DNA sequence will be passed on to *their* children, and their children's children, and so on. Through a process called neutral genetic drift, given enough time (it happens faster in smaller populations than large) the "fossil" viral DNA will either be flushed out of the population eventually, *or* by luck of the draw end up in every member of the population X generations down the road. It all depends on a roll of the genetic dice.
Due to the hurdles, "fossil" retroviral DNA strings (known by the technical name of "endogenous retroviruses") don't end up ubiquitous in a species very often, but it provably *does* happen. In fact, the Human DNA project has identified literally *thousands* of such fossilized "relics" of long-ago ancestral infections in the human DNA.
And several features of these DNA relics can be used to demonstrate common descent, including their *location*. The reason is that retroviruses aren't picky about where their DNA gets inserted into the host DNA. Even in an infection in a *single* individual, each infected cell has the retroviral DNA inserted into different locations than any other cell. Because the host DNA is so enormous (billions of basepairs in humans, for example), the odds of any retroviral insertion event matching the insertion location of any other insertion event are astronomically low. The only plausible mechanism by which two individuals could have retroviral DNA inserted into exactly the same location in their respective DNAs is if they inherited copies of that DNA from the same source -- a common ancestor.
Thus, shared endogenous retroviruses between, say, ape and man is almost irrefutable evidence that they descended from a common ancestor. *Unless* you want to suggest that they were created separately, and then a virus they were both susceptible to infected both a man and an ape in EXACTLY the same location in their DNAs (the odds of such a match by luck are literally on the order of 1,000,000,000,000,000,000 to 1...), *and* that the infections both happened in their gametocyte cells (combined odds on the order of 1,000,000 to 1) *and* that the one particular affected gametocyte is the one which produces the egg or sperm which is destined to produce an offspring (*HUGE* odds against), and *then* the resulting modified genome of the offspring becomes "fixed" in each respective population (1 out of population_size^squared)...
Then repeat that for *each* shared endogenous retrovirus (there are many) you'd like to claim was acquired independently and *not* from a shared ancestor...
Finally, you'd have to explain why, for say species A, B, and C, the pattern of shared same-location retroviruses is always *nested*, never *overlapped*. For example, all three will share some retroviruses, then A and B will both share several more, but if so then B *never* shares one with C that A doesn't also have (or at least remnants of).
In your "shared infection due to genetic similarities" suggestion, even leaving aside the near statistical impossibility of the infections leaving genetic "scars" in *exactly* the same locations in independent infections, one would expect to find cases of three species X, Y, and Z, where the degree of similarity was such that Y was "between" X and Z on some similarity scale, causing the same disease to befall X and Y but not Z, and another disease to affect Y and Z but not X. And yet, we don't find this in genetic markers. The markers are found in nested sequence, which is precisely what we would expect to see in cases of inheritance from common ancestry.
Here, for example, is an ancestry tree showing the pattern of shared same-location endogenous retroviruses of type HERV-K among primates:
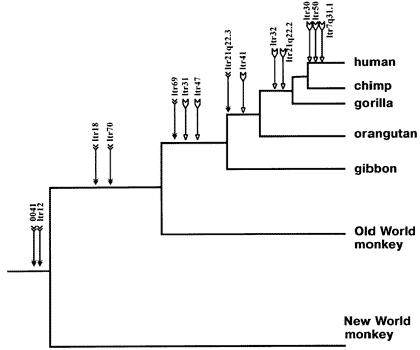
This is just a partial list for illustration purposes -- there are many more.
Each labeled arrow on the chart shows an ERV shared in common by all the branches to the right, and *not* the branches that are "left-and-down". This is the pattern that common descent would make. And common descent is the *only* plausible explanation for it. Furthermore, similar findings tie together larger mammal groups into successively larger "superfamilies" of creatures all descended from a common ancestor.
Any presumption of independent acquisition is literally astronomically unlikely. And "God chose to put broken relics of viral infections that never actually happened into our DNA and line them up only in patterns that would provide incredibly strong evidence of common descent which hadn't actually happened" just strains credulity (not to mention would raise troubling questions about God's motives for such a misleading act).
Once again, the evidence for common descent -- as opposed to any other conceivable alternative explanation -- is clear and overwhelming.
Wait, want more? Endogenous retroviruses are just *one* type of genetic "tag" that makes perfect sense evolutionary and *no* sense under any other scenario. In addition to ERV's, there are also similar arguments for the patterns across species of Protein functional redundancies, DNA coding redundancies, shared Processed pseudogenes, shared Transposons (including *several* independent varieties, such as SINEs and LINEs), shared redundant pseudogenes, etc. etc. Here, for example, is a small map of shared SINE events among various mammal groups:
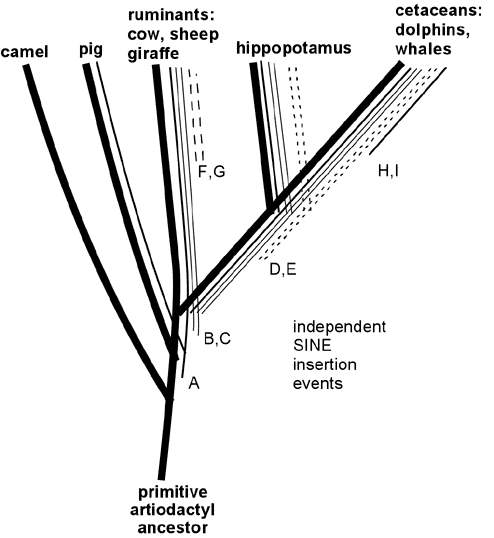
Like ERV's, any scenario which suggests that these shared DNA features were acquired separately strains the laws of probability beyond the breaking point, but they make perfect sense from an evolutionary common-descent scenario. In the above data, it is clear that the only logical conclusion is that, for example, the cetaceans, hippos, and ruminants shared a common ancestor, in which SINE events B and C entered its DNA and then was passed on to its descendants, yet this occurred after the point in time where an earlier common ancestor had given rise both to that species, and to the lineage which later became pigs.
And this pattern (giving the *same* results) is repeated over and over and over again when various kinds of molecular evidence from DNA is examined in detail.
The molecular evidence for evolution and common descent is overwhelming. The only alternative is for creationists to deny the obvious and say, "well maybe God decided to set up all DNA in *only* ways that were consistent with an evolutionary result even though He'd have a lot more options open to him, even including parts which by every measure are useless and exactly mimic copy errors, ancient infections, stutters, and other garbage inherited from nonexistent shared ancestors"...
Or how about:
Humans have 23 pairs of chromosomes ---chimps and gorillas have 24 pairs. How many pairs of chromosomes did the "common ancestor" have? Was it 23 or 24 pairs? How do you "evolve" missing or added chromosomes ---that would happen all at one time. The common ancestor had 24 chromosomes.
If you look at the gene sequences, you'll find that Chromosome 2 in humans is pretty much just 2 shorter chimpanzee chromosomes pasted end-to-end, with perhaps a slight bit of lost overlap:

(H=Human, C=Chimpanzee, G=Gorilla, O=Orangutan)
Somewhere along the line, after humans split off from the other great apes, or during the split itself, there was an accidental fusion of two chromosomes, end-to-end. Where there used to be 24 chromosomes, now there were 23, but containing the same total genes, so other than a "repackaging", the DNA "instructions" remained the same.
If a chimpanzee gives birth to a creature with 23 chromosomes, that offspring isn't going to be a well-formed chimpanzee able to survive well.
It is if the same genes are present, which they would be in the case of a chromosome fusion.
Evolve would imply the genetic material changes little by little --not some big loss of two chromosomes at once but I don't see how they'd go away gene by gene.
Tacking two chromosomes together end-to-end is not a "big loss" of genes, and it really is a "little by little" change in the total genetic code. It's just been "regrouped" a bit. Instead of coming in 24 "packages", it's now contained in 23, but the contents are the same.
So how, you might ask, would the chromosomes from the first 23-chromosome "fused" individual match up with the 24 chromosomes from its mate when it tried to produce offspring? Very well, thanks for asking. The "top half" of the new extra-long Chromosome 2 would adhere to the original chromosome (call it "2p") from which it was formed, and likewise for the "bottom half" which would adhere to the other original shorter chromosome (call it "2q"). In the picture above, imagine the two chimp chromosomes sliding over to "match up" against the human chromosome. The chimp chromosomes would end up butting ends with each other, or slightly overlapping in a "kink", but chromosomes have overcome worse mismatches (just consider the XY pair in every human male -- the X and the Y chromosome are *very* different in shape, length, and structure, but they still pair up).
In fact, the "rubbing ends" of the matched-up chimp chromosomes, adhering to the double-long human-type chromosome, would be more likely to become fused together themselves.
For studies in which recent chromosome fusions have been discovered and found not to cause infertility, see:
Chromosomal heterozygosity and fertility in house mice (Mus musculus domesticus) from Northern Italy. Hauffe HC, Searle JB Department of Zoology, University of Oxford, Oxford OX1 3PS, United Kingdom. hauffe@novanet.it An observed chromosome fusion: Hereditas 1998;129(2):177-80 A new centric fusion translocation in cattle: rob (13;19). Molteni L, De Giovanni-Macchi A, Succi G, Cremonesi F, Stacchezzini S, Di Meo GP, Iannuzzi L Institute of Animal Husbandry, Faculty of Agricultural Science, Milan, Italy.
J Reprod Fertil 1979 Nov;57(2):363-75 Cytogenetics and reproduction of sheep with multiple centric fusions (Robertsonian translocations). Bruere AN, Ellis PM
J Reprod Fertil Suppl 1975 Oct;(23):356-70 Cytogenetic studies of three equine hybrids. Chandley AC, Short RV, Allen WR.
In that last reference, the Przewalski horse, which has 33 chromosomes, and the domestic horse, with 32 chromosomes (due to a fusion), are able to mate and produce fertile offspring. Meanwhile, the question may be asked, how do we know that the human Chromosome 2 is actually the result of a chromsome fusion at/since a common ancestor, and not simply a matter of "different design"?
Well, if two chromsomes accidentally merged, there should be molecular remnants of the original chromosomal structures (while a chromosome designed from scratch would have no need for such leftover "train-wreck" pieces).
Ends of chromosomes have characteristic DNA base-pair sequences called "telomeres". And there are indeed remnants of telomeres at the point of presumed fusion on human Chromosome 2 (i.e., where the two ancestral ape chromosomes merged end-to-end). If I may crib from a web page:
Telomeres in humans have been shown to consist of head to tail repeats of the bases 5'TTAGGG running toward the end of the chromosome. Furthermore, there is a characteristic pattern of the base pairs in what is called the pre-telomeric region, the region just before the telomere. When the vicinity of chromosome 2 where the fusion is expected to occur (based on comparison to chimp chromosomes 2p and 2q) is examined, we see first sequences that are characteristic of the pre-telomeric region, then a section of telomeric sequences, and then another section of pre-telomeric sequences. Furthermore, in the telomeric section, it is observed that there is a point where instead of being arranged head to tail, the telomeric repeats suddenly reverse direction - becoming (CCCTAA)3' instead of 5'(TTAGGG), and the second pre-telomeric section is also the reverse of the first telomeric section. This pattern is precisely as predicted by a telomere to telomere fusion of the chimpanzee (ancestor) 2p and 2q chromosomes, and in precisely the expected location. Note that the CCCTAA sequence is the reversed complement of TTAGGG (C pairs with G, and T pairs with A).
Another piece of evidence is that if human Chromosome 2 had formed by chromosome fusion in an ancestor instead of being designed "as is", it should have evidence of 2 centromeres (the "pinched waist" in the picture above -- chromosomes have centromeres to aid in cell division). A "designed" chromosome would need only 1 centromere. An accidentally "merged" chromosome would show evidence of the 2 centromeres from the two chromosomes it merged from (one from each). And indeed, as documented in (Avarello R, Pedicini A, Caiulo A, Zuffardi O, Fraccaro M, Evidence for an ancestral alphoid domain on the long arm of human chromosome 2. Hum Genet 1992 May;89(2):247-9), the functional centromere found on human Chromosome 2 lines up with the centromere of the chimp 2p chromosome, while there are non-functional remnants of the chimp 2q centromere at the expected location on the human chromosome. As an aside, the next time some creationist claims that there is "no evidence" for common ancestry or evolution, keep in mind that the sort of detailed "detective story" discussed above is repeated literally COUNTLESS times in the ordinary pursuit of scientific research and examination of biological and other types of evidence. Common ancestry and evolution is confirmed in bit and little ways over and over and over again. It's not just something that a couple of whacky anti-religionists dream up out of thin air and promulgate for no reason, as the creationists would have you believe.
And:
[The poster known as Mr. LLLICHY wrote:] Here is that Vitamin C dataAfter discovering this same data on another thread along with more discussion than has appeared here (I've taken the liberty of pinging the participants of that discussion), I see what the "mystery" is supposed to be -- it's supposed be why did some sites have multiple mutations while (small) stretches of other sites had none? In other words, why do the mutations appear clustered?
(You know, it would really help if people explained their points and questions in more detail, instead of leaving people to guess what the poster was thinking...)
[LLLICHY wrote:] "U238" that decays thrice, pretty good trick when there is "U238" that does not decay at all in 50,000,000 years.
Actually, no site had mutations "thrice". Three different bases at a given site is only *two* mutations (one original base, plus two mutations from it to something else).
Here's the "mutation map" from the actual DNA data:
--1-12--1-1-1-1--------1112112--1---1-11-1--------1 ALL/n
No mutations ("-") in about half the sites, one mutation at several (17) sites, two mutations at three sites. The first thing to keep in mind that random processes tend to "cluster" more than people expect anyway. People expect "randomness" to "spread out" somewhat evenly, but instead it's usually more "clumped", for statistical reasons that would be a diversion to go into right now. So "that looks uneven" isn't always a good indication that something truly is non-random.
If you don't believe me on that, I wrote a program which made 23 mutations totally at random on a 51-site sequence, then repeated the process to see what different random outcomes would look like:
10 X$=STRING$(51,"-")
20 FOR I=1 TO 23
30 J%=INT(RND*51)+1
40 C$=MID$(X$,J%,1)
50 IF C$="-" THEN MID$(X$,J%,1)="1" ELSE MID$(X$,J%,1)=CHR$(ASC(C$)+1)
60 NEXT I
70 PRINT X$
80 GOTO 10
Yeah, it's BASIC, so sue me. Here's a typical screenful of the results: -21---1---2---111----2-----2-1121-------1---1--11-1
-1--1--21-11---1-1--1-1---1----1---21-11111---11---
3-11---3-----1-----11-2-1---1--1----3--2---1--1----
---1-1--22--1-1--2-2111--1-1111---1------1-------1-
---32----1-11-1-----1---2-231----1------1-----11--1
----2---21--1---4----1-------------11-1--111-11-211
11--1-1---1-----1--1------1----3111--1----111-2-1-2
1112---1-3-1----1-1-----1-1------121--111-------1-1
-111121--1----1----1-1-1-1-11-2---1-1-------1-111--
-----------11-1---11-11--------21----12211--1---131
--1-211-1-1----21--11-1-2----1--1----11---11-----11
12---1-13------------2---21-21---11-1-1-1--2-------
-----2-1---1-1----21--11-11-1---111-1--111-----2--1
-----1-----1-1-1-1---1-2----11-21-11--1-111---1-21-
---11--1-1-122-1-1-1--1-----2-1-1-1-------1-1---111
--2--11----2--1---12-2----1-1---1-1--1--12----1-1-1
-111-1-----1-1----------1-21111--1-2-11-11-1----11-
11-1--211-1221-----1--1-----11--1-2-1----------11--
-----1-12-11---2-1---11--1-2--1----11---111-1----11
11----1--12---12----1---31---1-11----2--1-11-1-----
---1--111-1--1-1-111----1-21----1-1-3---1------2--1
-2-11----1-1------1------2-1-1--111-111-1-1----1111
1--1--1-1---1-111111--2--1-1------112----2---11----
Notice how oddly "clustered" most of them look, including one run which left a 13-site stretch "absolutely untouched", contrary to intuition (while having *4* mutations at a single site!) Frankly, I don't see anything in the real-life DNA mutation map which looks any different from these truly random runs. Random events tend to cluster more than people expect. That solves the "mystery" right there.
Also, there may be a selection factor -- the GLO gene is a *lot* bigger than this. One has to wonder if this small 51-bp section was presented just because it was the one that looked "least random". That would be a no-no, since one can always hand-select the most deviant subset out of larger sample in order to artificially skew the picture.
However, since there are some interesting evolutionary observations to be made, let's look at that DNA data again, slightly rearranged:
TAC CCC GTG GAG GTG CGC TTC ACT CGG GCG GAC GAC ATC CTG CTG AGC CCC PIG
TAC CCC GTG GAG GTA CGC TTC ACT CGC GGG GAC GAC ATC CTG CTG AGC CCC BOS
TAC CCC GTA GAG GTG CGC TTC ACC CGA GGC GAT GAC ATT CTG CTG AGC CCC RAT
TAC CCC GTG GAG GTG CGC TTC ACC CGA GGT GAT GAC ATC CTG CTG AGC CCG MOUSE
TAC CCT GTG GGG GTG CGC TTC ACC CGG GGG GAC GAC ATC CTG CTG AGC CCC GUIN PIG
TAC CTG GTG GGG GTA CGC TTC ACC TGG AG* GAT GAC ATC CTA CTG AGC CCC HUMAN
TAC CTG GTG GGG CTA CGC TTC ACC TGG AG* GAT GAC ATC CTA CTG AGC CCC CHIMPANZEE
TAC CCG GTG GGG GTG CGC TTC ACC CAG AG* GAT GAC GTC CTA CTG AGC CCC ORANGUTAN
TAA CCG GTG GGG GTG CGC TTC ACC CAA GG* GAT GAC ATC ATA CTG AGC CCC MACAQUE
Here I've put spaces between codons, and clustered the closely-related species together: pig/cow as ungulates, rat/mouse for their obvious relationship, guinea pig right below them but separated because of the pseudogene nature of its GLO gene, then primates all in a group, with man's closest relative, the chimp, immediately below him, followed by the more distant orangutan, and the even more distant macaque. Also note that the top four have "working" GLO genes, and the bottom five have "broken" GLO pseudogenes. First, let's consider just the four species with working GLO genes. Evolution predicts that even over large periods of time, these genes will be "highly conserved", with natural selection weeding out mutations that could "break" the gene. Note that the mutations will still have occurred in individuals of the population, but natural selection will "discourage" that mutation from spreading into the general population.
And before we go any further, let's talk about the "universal genetic code". In all mammals (indeed, in almost all living organisms), each triplet of DNA sites cause a particular amino acid to be formed. The mapping of triplets (called "codons") to amino acids is as follows:
|
|
|
Second Position of Codon |
|
|
|
|
|
T |
C |
A |
G |
|
|
F
i
r
s
t
P
o
s
i
t
i
o
n |
T |
| TTT |
Phe |
[F] |
| TTC |
Phe |
[F] |
| TTA |
Leu |
[L] |
| TTG |
Leu |
[L] |
|
| TCT |
Ser |
[S] |
| TCC |
Ser |
[S] |
| TCA |
Ser |
[S] |
| TCG |
Ser |
[S] |
|
| TAT |
Tyr |
[Y] |
| TAC |
Tyr |
[Y] |
| TAA |
Ter |
[end] |
| TAG |
Ter |
[end] |
|
| TGT |
Cys |
[C] |
| TGC |
Cys |
[C] |
| TGA |
Ter |
[end] |
| TGG |
Trp |
[W] |
|
|
T
h
i
r
d
P
o
s
i
t
i
o
n |
| C |
| CTT |
Leu |
[L] |
| CTC |
Leu |
[L] |
| CTA |
Leu |
[L] |
| CTG |
Leu |
[L] |
|
| CCT |
Pro |
[P] |
| CCC |
Pro |
[P] |
| CCA |
Pro |
[P] |
| CCG |
Pro |
[P] |
|
| CAT |
His |
[H] |
| CAC |
His |
[H] |
| CAA |
Gln |
[Q] |
| CAG |
Gln |
[Q] |
|
| CGT |
Arg |
[R] |
| CGC |
Arg |
[R] |
| CGA |
Arg |
[R] |
| CGG |
Arg |
[R] |
|
|
| A |
| ATT |
Ile |
[I] |
| ATC |
Ile |
[I] |
| ATA |
Ile |
[I] |
| ATG |
Met |
[M] |
|
| ACT |
Thr |
[T] |
| ACC |
Thr |
[T] |
| ACA |
Thr |
[T] |
| ACG |
Thr |
[T] |
|
| AAT |
Asn |
[N] |
| AAC |
Asn |
[N] |
| AAA |
Lys |
[K] |
| AAG |
Lys |
[K] |
|
| AGT |
Ser |
[S] |
| AGC |
Ser |
[S] |
| AGA |
Arg |
[R] |
| AGG |
Arg |
[R] |
|
|
| G |
| GTT |
Val |
[V] |
| GTC |
Val |
[V] |
| GTA |
Val |
[V] |
| GTG |
Val |
[V] |
|
| GCT |
Ala |
[A] |
| GCC |
Ala |
[A] |
| GCA |
Ala |
[A] |
| GCG |
Ala |
[A] |
|
| GAT |
Asp |
[D] |
| GAC |
Asp |
[D] |
| GAA |
Glu |
[E] |
| GAG |
Glu |
[E] |
|
| GGT |
Gly |
[G] |
| GGC |
Gly |
[G] |
| GGA |
Gly |
[G] |
| GGG |
Gly |
[G] |
|
|
(The above table imported from http://psyche.uthct.edu/shaun/SBlack/geneticd.html, which also has a nice introduction to the genetic code.)
Another version of the same table with nifty Java features and DNA database lookups can be found here.
The thing which is most relevant to the following discussion is the fact that most of the genetic codes are "redundant" -- more than one codon (triplet) encodes to exactly the same amino acid. This means that even in genes which are required for the organism, certain basepair mutations make absolutely no difference if the change is from one codon which maps into amino acid X to another codon which still maps into amino acid X. (This fact allows certain kinds of evolutionary "tracers" to be "read" from the DNA, as described here).
Now back to our DNA data. The redundancy in the genetic code means that some basepair sites will have more "degrees of freedom" than others (i.e., ways in which they can mutate without disrupting the gene's biological function in any way). Let's look at the four species with working GLO genes again:
TAC CCC GTG GAG GTG CGC TTC ACT CGG GCG GAC GAC ATC CTG CTG AGC CCC PIG
TAC CCC GTG GAG GTA CGC TTC ACT CGC GGG GAC GAC ATC CTG CTG AGC CCC BOS
TAC CCC GTA GAG GTG CGC TTC ACC CGA GGC GAT GAC ATT CTG CTG AGC CCC RAT
TAC CCC GTG GAG GTG CGC TTC ACC CGA GGT GAT GAC ATC CTG CTG AGC CCG MOUSE
T T T A T A T T T A T C C T T T T T T T T
A A A A A C A A A A A
G C G G G G G C C C
--- --- --1 --- --1 --- --- --1 --2 -12 --1 --- --1 --- --- --- --1
Under each site of the mouse DNA, I've listed the "alternative" bases which could be be substituted for the mouse base at that site WITHOUT ALTERING THE GENE'S FUNCTION (because of genetic code redundancy). And under that I show the "mutation map" of just those four species. Note that most of the "alternative" bases are in the third base of each codon, *and* that this is where all but one of the mutations have appeared. This is because these were the sites which were "free" to mutate in the way they did, because the mutation was genetically neutral. That doesn't mean that the first and second sites of each codon were immune from mutation, it's just that when mutations did occur at those sites, natural selection weeded them out quickly because they most likely "broke" the GLO gene for the individuals which received that mutuation. What we see above is the results after natural selection has already "filtered" the undesirable mutations and left the ones which "do no harm".
Additionally, the two sites which have mutated twice (i.e. have a "2" in the mutation map) are ones which had more "allowable" mutations. Also note that the sites which had the fewest allowable alternatives (only one alternate letter allowed) didn't have any mutations fix at those sites, which is unsurprising since a "safe" mutation would be less likely to occur there versus a site that "allowed" two or three alternatives.
All this is as predicted by evolutionary theory, you'll note.
It also explains the one anomoly of the original mutation map, which is that the mutation counts do tend to be higher at the third base of a codon.
However... What about the one exception? The pig DNA has had one mutation at a site which does not encode to exactly the same amino acid (which is the case for *all* the other ones). In the pig DNA, the GGG codon (mapping to Glycine) has changed to a GCG codon (mapping to Alanine). What's up with that? Well, one of two things. First and most likely, just as base values in codons have a built-in redundancy, so do the amino acids which make up the proteins which result from the DNA templates. In other words, certain amino acids can be substituted for other ones at some sites in given proteins without making any functional difference. (This "protein functional redundancy" also has implications for "evolutionary tracer" analysis, see here.) That may well be the case for Alanine versus Glycine in the GLO protein, but I'm not enough of a biochemist to be able to say. The other option is that it *does* make some difference in the function of the pig GLO protein, but not enough to "break" the vitamin-C synthesis (as proven by the fact that pigs *can* synthesize vitamin C). So one way or another, it's not a deal-breaker even though pig GLO will not be 100% identical to cow/mouse/rat GLO. It's yet another "allowable" mutation.
More interesting evolutionary observations: The number of mutational differences between pig/cow is 3, the number between mouse/rat is 4, and the difference between rat/cow is 7 -- all roughly as one would expect from the evolutionary relatedness of these animals (cows/pigs and rats/mice are each closer to each other than the rodents are to the ungulates).
Now let's take a close look at the guinea pig:
TAC CCT GTG GGG GTG CGC TTC ACC CGG GGG GAC GAC ATC CTG CTG AGC CCC GUIN PIG
--- --1 --- -1- --- --- --- --- --1 --1 --1 --- --- --- --- --- ---
The "mutation map" under the guinea pig DNA is compared to the mouse DNA. Fascinating: Note that four of the five mutations are in the third base of a codon, *and* are of the type "allowed" by the genetic code redundancy. This indicates strongly that most of the evolutionary divergence between guinea pigs and mice likely occurred while the guinea pig's ancestors still had a working GLO gene. This is the sort of prediction implied by the evolutionary theory which could be cross-checked by further research of various types, and if verified, would be yet further confirmation that evolutionary theory is likely correct. So far, evolutionary theory has been subjected to literally countless tests like this, large and small, and the vast majority of results have confirmed the evolutionary prediction. This track record is hard to explain if evolution is an invalid theory, as some assert... Finally, let's look over the primate DNA and mutation map (relative to each other):
TAC CTG GTG GGG GTA CGC TTC ACC TGG AG* GAT GAC ATC CTA CTG AGC CCC HUMAN
TAC CTG GTG GGG CTA CGC TTC ACC TGG AG* GAT GAC ATC CTA CTG AGC CCC CHIMPANZEE
TAC CCG GTG GGG GTG CGC TTC ACC CAG AG* GAT GAC GTC CTA CTG AGC CCC ORANGUTAN
TAA CCG GTG GGG GTG CGC TTC ACC CAA GG* GAT GAC ATC ATA CTG AGC CCC MACAQUE
--1 -1- --- --- 1-1 --- --- --- 111 1-- --- --- 1-- 1-- --- --- ---
Evolutionary theory predicts that because the GLO gene is "broken" in primates (i.e. is a pseudogene), mutations in it are highly likely to be neutral (i.e., make no difference, since it can't get much more broken), and thus mutations are just as likely to accumulate at any site as any other. Is that what we see? Yup. There's no obvious pattern to the mutations between primates in the above mutation map, and unlike the pig/cow/mouse/rat mutation map, the mutations aren't predominantly at the "safer" third base of a codon, nor of a type that would be "safe". In fact, one base has vanished entirely, but no biggie, the gene's already broken. Also, although primates share a more recent common ancestor than cows/pigs/mice/rats, note that they've already racked up almost as many relative mutations as the cow/pig/mouse/rat DNA. This too is just as evolutionary theory predicts, because many mutations in a functional gene (GLO in this case) will be "non-safe" and weeded out by natural selection, making for a slower mutation fixation rate overall than in a pseudogene (as GLO is in primates) where natural selection doesn't "care" about the vast majority of mutations since *most* are neutral. So pseudogenes accumulate mutations faster than functional genes (even though rate of mutation *occurence* in both are likely the same).
Finally, note that there are ZERO mutational differences between the human DNA and the chimpanzee DNA, our nearest living relative.
I also see some interesting implications in the DNA sequences concerning which specific mutation fixed during what branch of the common-descent evolutionary tree for all the species represented, but reconstructing that would not only take another couple hours, at least, but would be a major bear to code in HTML, since I'd have to draw trees with annotations on the nodes... Bleugh.
In any case, I hope I've clarified some of the methods by which biologists find countless confirmations of evolution in DNA data. This is just a "baby" example, and to be more statistically valid would have to be done over much vaster sections of DNA sequences, but my intent was to demonstrate some of the concepts.
And if such a small amount of DNA as this can make small confirmations of evolutionary predictions, imagine the amount of confirmation from billion-basepair DNA data from each species compared across thousands of species... The amount of confirmatory discoveries for evolution from DNA analysis has already been vast, and promises to only grow in the future. For an overview of some of the different lines of evidence being studied, see The Journal of Molecular Evolution -- abstracts of all articles, current and back issues, can be browsed free online.
See also, for example (out of thousands):
Analysis of the human Alu Ye lineageHuman endogenous retrovirus HERV-K14 families: status, variants, evolution, and mobilization of other cellular sequences
Ancestral population sizes and species divergence times in the primate lineage on the basis of intron and BAC end sequences
Molecular and temporal characteristics of human retropseudogenes.
Evolution of RPS4Y
Inactivation of CMP-N-acetylneuraminic acid hydroxylase occurred prior to brain expansion during human evolution
Serine hydroxymethyltransferase pseudogene, SHMT-ps1: a unique genetic marker of the order primates
Structure and phylogenetic analysis of an endogenous retrovirus inserted into the human growth factor gene pleiotrophin
Molecular evolution of cytochrome c oxidase subunit IV: evidence for positive selection in simian primates
The ZNF75 zinc finger gene subfamily: isolation and mapping of the four members in humans and great apes
Insertions and duplications of mtDNA in the nuclear genomes of Old World monkeys and hominoids
Conservation of sequences between human and gorilla lineages: ADP-ribosyltransferase (NAD+) pseudogene 1 and neighboring retroposons
Fixation times of retroposons in the ribosomal DNA spacer of human and other primates
The gamma-globin genes and their flanking sequences in primates: findings with nucleotide sequences of capuchin monkey and tarsier
The emergence of new DNA repeats and the divergence of primates
Genetic diversity at class II DRB loci of the primate MHC
Nucleotide sequences of immunoglobulin-epsilon pseudogenes in man and apes and their phylogenetic relationships
Structure and evolution of human and African ape rDNA pseudogenes
How many more would you like?
Also, I can't quote the source, but I remember reading some time in the past year or so that Darwin returned to his "Christian" roots and rejected his earlier positions on his deathbed. (More food-for-thought-and-conversation!)
That's a myth, the so-called "Lady Hope story", concerning an alleged "deathbed confession" from Darwin.
It's not true, but even if it had been, it would mean exactly nothing. The validity of a scientific theory is based on the preponderance of evidence, confirmation of its predictions, and successfully passing potential falsification tests. It is not based on whether any particular person or scientist believes it, disbelieves it, repudiates it, etc. -- not even its founder. Evolution has passed the above hurdles with flying colors, and even if Darwin had chickened out long ago, it would remain just as validated by subsequent tests.
To: Ichneumon; Prov3456
Actually Itchy, we've moved on from Hovind. What my "problem, not Ken Ham's" is, is that I consider Ken Ham lacking in basic human decency - his views on evolution aside.
153
posted on
03/20/2005 3:08:45 PM PST
by
Oztrich Boy
(Rules are for the guidence of wise men, and the blind obedience of fools.)
To: Prov3456
"That, IMO, is the problem. If you'd argue that species change/mutate within themselves, then I'd agree. But it's the idea that monkeys BECAME people (ie. one kind of DNA turning into another kind of DNA) that disproves Darwin's theory for me (and, I suspect, more and more scientists as more is learned about biology, DNA, etc)."
First, let me say that the definition of the word "species" is a little "wishy washy" in my opinion. It is applied different ways at different times. Generally it refers to organisms that have become SO DIFFERENT, that they have lost the ability to reproduce between themselves. Sometimes this means that they are genetically incapable of producing offspring, sometimes it means that behaviorally they do not produce offspring. Point being, "species" can seem and often is, rather an arbitrary point on a continuum between point A and point B.
The donkey and horse are a good example of "borderline", they are similar enough to produce a mule when you cross them, but are different enough that the offspring is "not quite right", its sterile.
Species do not "pop" into existence (although some features might like new flower color for example), they are the result of generations of changes.
I'm forgetting the name of the species (I'll look it up if someone REALLY needs to know), but there is a bird, a type of seagull as I recall, that is found all the way around the world. If you compare one population to the population a little east of it, they are slightly different, but without doubt the same species. By the time you follow the loop all the way around the world and the extremes of the two ends meet, they are so different, that they do not interbreed.
If the intermediate populations were to die out (comet, volcano, disease, whatever), these would without doubt become considered two species. They are just NOT the same bird. It is only the existence of a continuum from point A to point B that allows these birds to be considered the same species.
154
posted on
03/20/2005 3:15:37 PM PST
by
ndt
To: Prov3456
Correct me if I'm wrong, but didn't Darwin's Origin of the Species postulate that through natural selection and mutation new species would be produced/created?New species have in fact been seen to evolve many times. What this means, applied to sexually reproducing organism, is that specific populations differentiate to the point where they can no longer exchange genes. And such speciation (species formation) has taken place both in the laboratory and in the wild, for instance with several kinds of fruit flies and other insects.
One problem in getting this fact across is that people often demand to be shown entirely novel kinds of organism appear before their very eyes. But it doesn't work that way. Even rapid speciation usually takes thousands or at the very least, hundreds of years, and especially when we are talking about megafauna like horses and humans. That is why we are normally confined to seeing evolution unfold in the fossil chain, which inevitably leaves some leeway for the willful disbelief of dogmatic creationists ("hey, that Brontosaurus is constructed out of chicken bones! It says so in this pamphlet!")
That, IMO, is the problem. If you'd argue that species change/mutate within themselves, then I'd agree. But it's the idea that monkeys BECAME people (ie. one kind of DNA turning into another kind of DNA) that disproves Darwin's theory for me (and, I suspect, more and more scientists as more is learned about biology, DNA, etc).
The kind of DNA does not need to change, nor does it in fact. Even the content is mostly the same among closely related species - famously, humans and chimpanzees have more than 98 percent of their DNA in common. Anyway, how does any of this 'disprove' the theory of evolution? The fact that some people profess to find it incredible is not a logical objection to the theory.
Also, I can't quote the source, but I remember reading some time in the past year or so that Darwin returned to his "Christian" roots and rejected his earlier positions on his deathbed.
That, my friend, has the dubious distinction of being both wrong and irrelevant. For one thing, the notion that Darwin retracted on his deathbed has been discredited. Its source is a certain religious lady claiming to have visited him on his deathbed; her story has been taken apart and has no independent support. For another thing, even if true this story would not make an iota of difference today. You see, in science it is evidence that matters, not personal authority. It could turn out that Einstein disbelieved his own Theory of Relativity and that would be make a dent in that theory's status as accepted science because it is borne out by multiple experiments and allows accurate predictions. However, it might in principle be refuted tomorrow, just as the theory of evolution would be by, say, the finding of a fossilized mammuth in a pre-cambrium rock formation. That's how intellectually honest creationists should go about it. But instead they prefer to simply make stuff up, and then whine about how they are 'oppressed' by actual scientists.
In fact, that lying lady with her deathbed retraction story is the perfect symbol of their lazy, dishonest ilk.
To: ndt; Prov3456
If the intermediate populations were to die out (comet, volcano, disease, whatever), these would without doubt become considered two species. They are just NOT the same bird. It is only the existence of a continuum from point A to point B that allows these birds to be considered the same species. Actually the two ends of the loop: The Herring Gull (Larus argentatus) and Lesser Black-backed Gull (L. fuscus), are considered to be two species, because that's how they are where their habitats overlap in the British Isles.
156
posted on
03/20/2005 3:27:14 PM PST
by
Oztrich Boy
(Rules are for the guidence of wise men, and the blind obedience of fools.)
To: Strategerist
Many of these theaters are in science museums and they have a responsbility to educate accurately. That's what's so disturbing about this. How many Luddites actually go to a museum?
157
posted on
03/20/2005 3:55:08 PM PST
by
balrog666
(A myth by any other name is still inane.)
To: Oztrich Boy
Thank you, I was too lazy to look up the name.
158
posted on
03/20/2005 4:03:10 PM PST
by
ndt
To: Ichneumon
159
posted on
03/20/2005 4:36:51 PM PST
by
PatrickHenry
(<-- Click on my name. The List-O-Links for evolution threads is at my freeper homepage.)
To: ndt
On the contrary, all it takes is one really good, reproducible experiment to blow a theory out of the water. Well evolution certainly can't produce such an experiment. This subject matter doesn't lend itself to such textbook experiments.
I don't have to produce anything. The evolutionists have yet to demonstrate any evidence that man evolved from single celled creatures by chance. Its just a theory. Another theory, and I believe to be more plausable, is that the complexity of man arose based on design by an intelligent creator. Both are just mere theories by mere men.
However, competing theories aren't even allowed in many classrooms because of the leftist NEA and the atheists. The evolutionists demand that their theories be presented as the other theory in the classroom. Its a load of crap.
Navigation: use the links below to view more comments.
first previous 1-20 ... 121-140, 141-160, 161-180 ... 221-236 next last
Disclaimer:
Opinions posted on Free Republic are those of the individual
posters and do not necessarily represent the opinion of Free Republic or its
management. All materials posted herein are protected by copyright law and the
exemption for fair use of copyrighted works.
FreeRepublic.com is powered by software copyright 2000-2008 John Robinson