Skip to comments.
Seattle think tank raises questions about evolution
Charlotte Observer & The Seattle Times ^
| 04/05/2005
| LINDA SHAW
Posted on 04/05/2005 7:42:56 AM PDT by bedolido
SEATTLE - (KRT) - Three years ago, the Ohio Board of Education invited a small but influential Seattle think tank to debate the way evolution is taught in Ohio schools.
It was an opportunity for the Discovery Institute to promote its notion of intelligent design, the controversial idea that parts of life are so complex they must have been designed by some intelligent agent.
Instead, leaders of the institute's Center for Science and Culture decided on what they consider a compromise. Forget intelligent design, they argued, with its theological implications. Just require teachers to discuss evidence that refutes Charles Darwin's theory of evolution, as well as what supports it.
They called it "teach the controversy," and that's become the institute's rallying cry as a leader in the latest efforts to raise doubts about Darwin in school. Evolution controversies are brewing in eight school districts, half a dozen state legislatures and three state boards of education, including the one in Kansas, which wrestled with the issue in 1999 as well.
"Why fight when you can have a fun discussion?" asks Stephen Meyer, the center's director. The teach-the-controversy approach, he said, avoids "unnecessary constitutional fights" over the separation of church and state, yet also avoids teaching Darwin's theories as dogma.
But what the center calls a compromise, most scientists call a creationist agenda that's couched in the language of science.
There is no significant controversy to teach, they say.
"You're lying to students if you tell them that scientists are debating whether evolution took place," said Eugenie Scott, director of the National Center for Science Education, a nonprofit group that defends teaching of evolution in school.
The Discovery Institute, she said, is leading a public-relations campaign, not a scientific endeavor.
The Discovery Institute is one of the leading organizations working nationally to change how evolution is taught. It works as an adviser, resource and sometimes a critic with those who have similar views.
"There are a hundred ways to get this wrong," said Meyer. "And only a few to get them right."
Ohio got it right, he said, when its state Board of Education voted in 2002 to require students to learn that scientists "continue to investigate and critically analyze aspects of evolutionary theory."
Scott said it was a small victory at most for intelligent-design supporters, but Meyer considers it a significant one - a model other states should follow. Minnesota has adopted similar language.
The School Board in Dover, Pa., however, got it wrong, Meyer said, when it required instruction in intelligent design. (The matter is now in court.) Intelligent design isn't established enough yet for that, Meyer said.
He also criticizes the Georgia school board that put stickers on biology textbooks with a surgeon-general-like warning that evolution is "a theory not a fact." The stickers were a "dumb idea," he said bluntly. (A Georgia court ruled they were illegal, and the case is under appeal.)
In Wisconsin, the institute hopes it helped the school board in the small town of Grantsburg switch to a teach-the-controversy approach.
In each place, the institute says it responded to requests for help, although it's working to become more proactive, too. Some critics suspect the ties are even closer.
The Center for Science and Culture opened in 1996 as a part of the already-established Discovery Institute, which also studies more earthbound topics such as transportation, economics, technology and bioethics.
Founder Bruce Chapman - who has worked as an official in the Reagan administration, head of the U.S. Census Bureau and Washington's secretary of state - became interested in intelligent design after reading a piece Meyer wrote for The Wall Street Journal.
Meyer, then a philosophy professor at Whitworth College in Spokane, Wash., was defending a California professor in trouble for talking about intelligent design in biology class. To Chapman, it was an issue of academic freedom.
He invited Meyer to come speak at the institute. The more they talked, the more Chapman and others at the institute became interested in offering a home to Meyer and others interested in intelligent design.
Intelligent design appealed to their view that life isn't really as unplanned or unguided as Darwin's theories can make it seem.
"It interested me because it seemed so different than the reductionist science that came out of the 19th century ... that everything could be reduced to chemistry," said John West, a political scientist and center associate director.
The private institute has an annual budget of about $3.2 million, and plans to spend about $1.3 million on the intelligent-design work, Chapman said, mostly to support the work of about three dozen fellows.
The Fieldstead Charitable Trust, run by Christian conservative Henry Ahmanson and his wife, is one of the largest donors to that effort. Chapman declines to name more.
Meyer, the center's director, is a tall, friendly man who has undergraduate degrees in geology and physics and a Ph.D. in the philosophy of science from Cambridge, where he wrote his doctorate on the origins of life.
He says he's no creationist. He doesn't, for example, believe in a literal reading of the Bible, which would mean the Earth is about 6,000 years old.
He doesn't dispute that natural selection played a role in evolution; he just doesn't think it explains everything.
He often points to the Cambrian Period, a time more than 500 million years ago when most of the major groups of animals first appear in the fossil record. Meyer and other Discovery Institute fellows say those groups show up too fast, geologically speaking, to have come about through natural selection. That's one of what they see as controversies they want taught in school.
Scientists, however, say the Cambrian Period may not be completely understood, but that doesn't mean the theory of evolution is in trouble.
"They harp and harp on natural selection, as if natural selection is the only thing that evolutionary biologists deal with," said Scott. "Who knows whether natural selection explains the Cambrian body plans. ... So what?"
Scientists consider Meyer a creationist because he maintains some unnamed intelligence - and Meyer said he personally thinks it is God - has an active hand in creating some complex parts of life.
"I don't know what else to call it other than creationism," said Michael Zimmerman, a critic and dean at the University of Wisconsin Oshkosh.
Meyer, however, said he's a scientist who starts with scientific evidence, not the Bible. His goal - a big one - is to change the very definition of science so that it doesn't rule out the possibility that an intelligent designer is actively at work.
"Science should be open to whatever cause ... can best explain the data," Meyer said.
That would be a major change for science, which limits itself to the natural world. Scott said it would be a "science stopper."
"Once you allow yourself to say God did it, you stop looking for naturalistic explanations. If you stop looking, you won't find them," she said.
Scott said science isn't an atheistic worldview. In science, she said, "It is equally inappropriate to say God did it, or God had nothing to do with it."
The institute's call to "teach the controversy" meets strong resistance.
"There's no controversy about whether living things have common ancestors," Scott said. "There's no controversy about whether natural selection is very important in creating the variety of organisms we have today."
While the institute touts its list of 370 scientists who have signed a statement saying they have some doubts about Darwin's theory of natural selection, Scott's organization, in a parody of that effort, has a list of 500 names limited to scientists named Steve or Stephanie, in honor of the late Stephen Jay Gould, a well-known biologist who once wrote that evolution is "one of the best documented, most compelling and exciting concepts in all of science."
Public opinion is mixed. Many Christian denominations, including Catholics, see no contradiction between evolution and their faith, but a Gallup Poll last November found that only about a third of the respondents think Darwin's theory of evolution is well supported by scientific evidence.
Meyer hopes the Kansas Board of Education will invite the center to speak at its hearings in May. Speakers will be asked to address the issue the center wants to highlight: whether Kansas' science curriculum helps students understand debate over controversial topics such as evolution.
Kansas Citizens for Science, however, has urged a boycott of the hearings, saying the proposals have been "rejected by the science community at large."
TOPICS: Culture/Society; Extended News; Miscellaneous; News/Current Events; Philosophy; US: Washington
KEYWORDS: crevolist; evolution; questions; seattle; tank; think
Navigation: use the links below to view more comments.
first previous 1-20 ... 101-120, 121-140, 141-160 ... 201-213 next last
To: dominic7
My point is that fittest and survival do not go hand in hand. We agree on that point. However, I'm still not sure whether "survival" or "fitness" have any meaning to evolution except insofar as they enable reproduction.
121
posted on
04/05/2005 11:29:55 AM PDT
by
r9etb
To: r9etb
Well, evolutionary theory CAN account for these observations. And the existence of human fetal tails actually supports "macro" evolution.
If ID attempts to be a valid alternative theory, then it is obligated to explain observed facts.
122
posted on
04/05/2005 11:30:50 AM PDT
by
thomaswest
(We are all for God. Who claims to know is questionable.)
To: DesertSapper
Nice try Radio. I don't buy it. You cannot have done much research and not known that a dog and a wolf are of the same species.Huh? Where did I make comments on Dogs?
I'll read your site (already am) and you spend some time looking at the other side too. That's all I ask of any evolutionist - especially if they are teaching our kids!
I already told you I read your links.
I'll thank God for creation (for the both of us) - then we can debate how everything got to this point. I'm not all that sure that this debate means much to Him as long as His handiwork is acknowledged.
Not science, but I thank you for the prayer. :-)
I do love this argument and will readily admit that I know very little in the scheme of things. However, I have been reading on both sides of this issue for a few years now and will continue to do so.
Cool. :-) I will continue to read and study as well.
I will let pass your comments about my homeschooling as I answer to the God on how I raise my kids - not you. But that is not the point here is it? The point is; should we continue to allow our kids to receive only one flawed origins theory and intentionally ignore others? Even if you agree with evolution, do you ignore alternate theories?
Where did I comment about homeschooling???
I do teach my boys about macro-evolution but do not promote it as I don't believe it. Why can I not expect at least as much from public schools I am funding?
You sure you are posting to the right person?
To: thomaswest
Well, evolutionary theory CAN account for these observations. Evolution can account for Great Big Meteors and Ice Ages, but a theory of intelligent design cannot? It's as I thought -- you're creating a straw man.
As for the tails -- you didn't answer my question.
124
posted on
04/05/2005 11:32:20 AM PDT
by
r9etb
To: bedolido
I live very near Seattle and when I read the words "Seattle Think Tank" I broke out laughing uproariously.
125
posted on
04/05/2005 11:32:22 AM PDT
by
trubluolyguy
("If the enemy is in range, so are you")
To: thomaswest
If the saber-toothed tiger was intelligently designed to survive in the designed environment, why did this kind become extinct? The Designer's younger and dumber Sister rearranged the climate while deciding what clothes to wear.
126
posted on
04/05/2005 11:36:33 AM PDT
by
Doctor Stochastic
(Vegetabilisch = chaotisch is der Charakter der Modernen. - Friedrich Schlegel)
To: Rebel_Ace
Suppose pixies from the netherworld created life. Suppose Mars is made of reddish colored styrofoam. (We have not retreived samples, we just have pictures) Suppose, suppose, suppose. You can suppose any half-assed idea, but it is NOT "good science" to be chasing down and refuting ideas for which there is NO physical evidence, or measurable phenomenon.
Um, the no physical evidence is the refutation. Lack of physical evidence is NOT a problem for ID. The complexity of the biosphere is physical evidence. Whether it is compelling physical evidence is another question entirely. But to say there is no physical evidence for intelligent design is to wilfully turn a blind eye on the biological world around us.
127
posted on
04/05/2005 11:37:35 AM PDT
by
frgoff
To: trubluolyguy
From what I understand about Seattle politics, the words "Seattle" and "think" should never be used in a sentence together ... lol
128
posted on
04/05/2005 11:37:47 AM PDT
by
dartuser
(Many people think that questioning Darwinian evolution must be equivalent to espousing creationism.)
To: r9etb
"Why should ID have to account for them?"
DING! DING! DING! DING!
Game Over, thanks for playing! See, when folks observe puzzling physical facts, the process of science tries to explain and account for them. That's what "good science" does. Theories and ideas are developed to explain observations, and attempt to make useful predictions about conditions not yet observed, and hopefully devise conclusive experiments to refine or reject the theory or model being considered.
While evolutionary theory might attempt to explain fetal developmental stages in terms of inherited, but now functionally obsolete genetic information, we see now that the ID theory does not need to even attempt to explain anything inconvenient.
Actually, this is a GREAT argument. I'll try to use it myself later.
Where did the petty cash go?
"Why should I have to account for it?"
Why does a Hot Air balloon rise?
"Why should physics have to account for that?"
Why do extraordinarily hairy men insist on wearing Speedos at the beach?
"Why should I have to account for them?"
OK, that last one was just silly, but I do wonder...
129
posted on
04/05/2005 11:39:02 AM PDT
by
Rebel_Ace
(Tags?!? Tags?!? We don' neeeed no stinkin' Tags!)
To: Wonder Warthog
There is no such distinction (i.e. there is no such theory as "micro-evolution"). The idea of "micro-evolution" is a weasel-word concept because the creationists CANNOT deny the facts of genetic change caused by the mutation/natural selection mechanism---so they try to have it both ways by coining new language.Actually, it's a distinction made based on observation. Evolutionists try to mash the two together in a classic post hoc ergo propter hoc fallacy.
130
posted on
04/05/2005 11:43:23 AM PDT
by
frgoff
To: LauraleeBraswell
Evolution- Like that some wolves evolved into dogs. That's proven.Correction. Wolves were bred into dogs.
131
posted on
04/05/2005 11:45:48 AM PDT
by
frgoff
To: dartuser; Rebel_Ace; DesertSapper; PatrickHenry
Good argument ... except evolutionists do exactly the same thing yet are not criticized for it. Recall the net theory of how birds first took to powered flight. They were better than their ancestors because they could use their wings to catch more bugs. And by flapping to catch the bugs they eventually took off and flew. Now what empirical evidence was there for this theory of flight? There was none, nor could there be. What have you been smoking? Contrary to your dishonest implication, that evolutionary scenario was not just pulled out of thin air without any evidence -- instead, it was developed as a direct result of examining the vast amounts of evidence and determining what the evidence indicated. The fossil evidence on this matter is clear.
What creationist source did you get these falsehoods from?
From a previous post of mine:
The cladogram for the evolution of flight looks like this:

(Note -- each name along the top is a known transitional fossil; and those aren't all that have been discovered.) Here's a more detailed look at the middle section:

Fossils discovered in the past ten years in China have answered most of the "which came first" questions about the evolution of birds from dinosaurs.
We now know that downy feathers came first, as seen in this fossil of Sinosauropteryx:

That's a close-up of downy plumage along the backbone. Here's a shot of an entire fossil

Sinosauropteryx was reptilian in every way, not counting the feathers. It had short forelimbs, and the feathers were all the same size. Presumably, the downy feathers evolved from scales driven by a need for bodily insulation.
Next came Protarchaeopteryx:

It had long arms, broad "hands", and long claws:

Apparently this species was driven by selection to develop more efficient limbs for grasping prey. One of the interesting things about this species is that the structure of the forelimb has been refined to be quite efficient at sweeping out quickly to grab prey, snap the hands together, then draw them back towards the body (mouth?). The specific structures in question are the semilunate carpal (a wrist bone), that moves with the hand in a broad, flat, 190 degree arc, heavy chest muscles, bones of the arm which link together with the wrist so as to force the grasping hands to spread out toward the prey during the forestroke and fold in on the prey during the upstroke. Not only is this a marvelously efficient prey-grabbing mechanism, but the same mechanism is at the root of the wing flight-stroke of modern birds. Evolution often ends up developing a structure to serve one need, then finds it suitable for adaptation to another. Here, a prey-grasping motion similar in concept to the strike of a praying mantis in a reptile becomes suitable for modifying into a flapping flight motion.
Additionally, the feathers on the hands and tail have elongated, becoming better suited for helping to sweep prey into the hands.
Next is Caudipteryx:

This species had hand and tail feathers even more developed than the previous species, and longer feathers, more like that of modern birds:

However, it is clear that this was still not a free-flying animal yet, because the forelimbs were too short and the feathers not long enough to support its weight, and the feathers were symmetrical (equal sized "fins" on each side of the central quill). It also had very reduced teeth compared to earlier specimens and a stubby beak:

But the elongation of the feathers indicates some aerodynamic purpose, presumably gliding after leaping (or falling) from trees which it had climbed with its clawed limbs, in the manner of a flying squirrel. Feathers which were developed "for" heat retention and then pressed into service to help scoop prey were now "found" to be useful for breaking falls or gliding to cover distance (or swooping down on prey?).
Next is Sinornithosaurus:

Similar to the preceding species, except that the pubis bone has now shifted to point to the back instead of the front, a key feature in modern birds (when compared to the forward-facing publis bone in reptiles). Here are some of the forearm feathers in detail:

Long feathers in detail:

Artists' reconstruction:

Next is Archaeopteryx:

The transition to flight is now well underway. Archaeopteryx has the reversed hallux (thumb) characteristic of modern birds, and fully developed feathers of the type used for flight (long, aligned with each other, and assymetrical indicating that the feathers have been refined to function aerodynamically). The feathers and limbs are easily long enough to support the weight of this species in flight. However, it lacks some structures which would make endurance flying more practical (such as a keeled sternum for efficient anchoring of the pectoral muscles which power the downstroke) and fused chest vertebrae. Archaeopteryx also retains a number of clearly reptilian features still, including a clawed "hand" emerging from the wings, small reptilian teeth, and a long bony tail. After the previous species' gliding abilities gave it an advantage, evolution would have strongly selected for more improvements in "flying" ability, pushing the species towards something more resembling sustained powered flight.
Next is Confuciusornis:

This species had a nearly modern flight apparatus. It also displays transitional traits between a reptilian grasping "hand" and a fully formed wing as in modern birds -- the outer two digits (the earlier species had three-fingered "hands") in Confuciusornis are still free, but the center digit has now formed flat, broad bones as seen in the wings of modern birds.

Additionally, the foot is now well on its way towards being a perching foot as in modern birds:

It also has a keeled sternum better suited for long flight, and a reduced number of vertebrae in the tail, on its way towards becoming the truncated tail of modern birds (which while prominent, is a small flap of muscle made to look large only because of the long feathers attached).
From this species it's only a small number of minor changes to finish the transition into the modern bird family.
(Hey, who said there are no transitional fossils? Oh, right, a lot of dishonest creationists. And there are a lot more than this, I've just posted some of the more significant milestones.)
There's been a very recent fossil find along this same lineage, too new for me to have found any online images to include in this article. And analysis is still underway to determine exactly where it fits into the above lineage. But it has well-formed feathers, which extend out from both the "arms" and the legs. Although it wasn't advanced enough to fully fly, the balanced feathering on the front and back would have made it ideally suited for gliding like a flying squirrel, and it may be another link between the stage where feathers had not yet been pressed into service as aerodynamic aids, and the time when they began to be used more and more to catch the air and developing towards a "forelimbs as wings" specialization.
So in short, to answer your question about how flight could have developed in birds, the progression is most likely some minor refinement on the following:
1. Scales modified into downy feathers for heat retention.
2. Downy feathers modified into "straight" feathers for better heat retention (modern birds still use their body "contour feathers" in this fashion).
3. Straight feathers modified into a "grasping basket" on the hands (with an accompanying increase in reach for the same purpose).
4. Long limbs with long feathers refined to better survive falls to the ground.
5. "Parachute" feathers refined for better control, leading to gliding.
6. Gliding refined into better controlled, longer gliding.
7. Long gliding refined into short powered "hops".
8. Short powered flight refined into longer powered flight.
9. Longer powered flight refined into long-distance flying.
Note that in each stage, the current configuration has already set the stage for natural selection to "prefer" individuals which better meet the requirements of the next stage. Evolution most often works like this; by taking some pre-existing ability or structure, and finding a better use for it or a better way to make it perform its current use.
To: r9etb
As requested. See references imbedded.
http://www.talkorigins.org/faqs/comdesc/section2.html#ontogeny_ex4
Example 4: the embryonic human tail
Humans are classified by taxonomists as apes; one of the defining derived characters of apes is the lack of an external tail. However, human embryos initially develop tails in development. At between four and five weeks of age, the normal human embryo has 10-12 developing tail vertebrae which extend beyond the anus and legs, accounting for more than 10% of the length of the embryo (Fallon and Simandl 1978; Moore and Persaud 1998, pp. 91-100; Nievelstein et al. 1993). The embryonic tail is composed of several complex tissues besides the developing vertebrae, including a secondary neural tube (spinal cord), a notochord, mesenchyme, and tail gut. By the eighth week of gestation, the sixth to twelfth vertebrae have disappeared via cell death, and the fifth and fourth tail vertebrae are still being reduced. Likewise, the associated tail tissues also undergo cell death and regress.
Figure 2.4.3. A histological cross-section of the human embryo's tail at Carnegie stage 14. The complex structures present in the human tail were visualized with light microscopy in this image. At Carnegie stage 14 (about 32 days old), the human tail is composed of neural tube (n, doubled in a large fraction of embryos), notochord (c), developing vertebrae (somites, s), gut (g), and mesenchyme (m). These specialized structures extend for the length of the tail, and the cells of all these structures die and are digested by immune system macrophages within the next two weeks of embryonic development.
...... In adult humans, the tail is finally reduced to a small bone composed of just four fused vertebrae (the coccyx) which do not protrude from the back (Fallon and Simandl 1978; Sapunar et al. 2001) (see Figure 2.4.1).
The regression of the human embryonic tail can be clearly seen in the fantastic images available at the Multi-dimensional Human Embryo site, where online images of three-dimensional MRI scans of live human embryos are archived. Different levels of maturity of the human embryo are classified according to the Carnegie stages. The embryonic post-anal tail is clearly visible in Carnegie stages 14, 15, and 16. The site has movies of a human embryo in rotation, giving clear views of the embryo's three-dimensional contours. Most stages have movies with the neural tube highlighted. It is especially informative to compare these rotating movies of the early stages (e.g. Carnegie stage 14 or stage 15) with the last stage (Carnegie stage 23), where the regression by cell death of the neural tube in the tail is clearly evident.
133
posted on
04/05/2005 11:48:55 AM PDT
by
thomaswest
(We are all for God. Who claims to know is questionable.)
To: Doctor Stochastic
Was that before or after Noah's Flood?
Or, why weren't passenger pigeons better designed to avoid buckshot?
134
posted on
04/05/2005 11:52:10 AM PDT
by
thomaswest
(We are all for God. Who claims to know is questionable.)
To: thomaswest
And the existence of human fetal tails actually supports "macro" evolution. Octogeny recapitulating Phylogeny has been discredited for probably over thirty years, now.
135
posted on
04/05/2005 12:02:47 PM PDT
by
frgoff
To: Rebel_Ace
Game Over, thanks for playing! See, when folks observe puzzling physical facts, the process of science tries to explain and account for them. DUNG! DUNG! DUNG! DUNG!
You clearly don't understand the point being made, which is that there is no more reason for ID to account for Mass Extinctions (currently thought to result from things like Giant Meteor Impacts) than for the theory of evolution to do so. Giant Meteors are explicitly outside the context of both concepts: organisms will be killed by them regardless of whether they were evolved, designed, or some combination of the two.
Get it?
136
posted on
04/05/2005 12:09:51 PM PDT
by
r9etb
To: bedolido
"Why fight when you can have a fun discussion?" asks Stephen Meyer, the center's director. The teach-the-controversy approach, he said, avoids "unnecessary constitutional fights" over the separation of church and state, yet also avoids teaching Darwin's theories as dogma. Since Dembski, Behe and others have already admitted that evolution occurs, what's the controversy?
To: DesertSapper; RadioAstronomer
What? Just links? You'd rather we post a few hundred megabytes of evidence right here in the thread? Don't be silly.
Come on, I'm all for doing my research but don't you think you could at least pull some pertinate info OUT of this site for us?
That link *was* to the pertinant info regarding your specific question. And it was a brief overview at that.
Assume I'm lazy
You want us to believe that you're too lazy to click a link, but not to lazy to read the same material if we post it within the thread? Ooookay.
or maybe it's just that I don't have time - at the moment - to read yet another pro-evolution website.
How convenient.
This one is new to me though and I have begun to read it (and put it on my "favorites" for now).
That's a start, anyway.
But since you insist on material inline to the thread, here are some prior posts of mine that address your point:
Background: Retroviruses reproduce by entering a cell of a host (like, say, a human), then embedding their own viral DNA into the cell's own DNA, which has the effect of adding a "recipe" for manufacturing more viruses to the cell's "instruction book". The cell then follows those instructions because it has no reason (or way) to "mistrust" the DNA instructions it contains. So the virus has converted the cell into a virus factory, and the new viruses leave the cell, and go find more cells to infect, etc.

However, every once in a while a virus's invasion plans don't function exactly as they should, and the virus's DNA (or portions of it) gets embedded into the cell's DNA in a "broken" manner. It's stuck into there, becoming part of the cell's DNA, but it's unable to produce new viruses. So there it remains, causing no harm. If this happens in a regular body cell, it just remains there for life as a "fossil" of the past infection and goes to the grave with the individual it's stuck in. All of us almost certainly contain countless such relics of the past viral infections we've fought off.
However... By chance this sometimes happens to a special cell in the body, a gametocyte cell that's one of the ones responsible for making sperm in males and egg cells in females, and if so subsequent sperm/eggs produced by that cell will contain copies of the "fossil" virus, since now it's just a portion of the entire DNA package of the cell. And once in a blue moon such a sperm or egg is lucky enough to be one of the few which participate in fertilization and are used to produce a child -- who will now inherit copies of the "fossilized" viral DNA in every cell of his/her body, since all are copied from the DNA of the original modified sperm/egg.
So now the "fossilized" viral DNA sequence will be passed on to *their* children, and their children's children, and so on. Through a process called neutral genetic drift, given enough time (it happens faster in smaller populations than large) the "fossil" viral DNA will either be flushed out of the population eventually, *or* by luck of the draw end up in every member of the population X generations down the road. It all depends on a roll of the genetic dice.
Due to the hurdles, "fossil" retroviral DNA strings (known by the technical name of "endogenous retroviruses") don't end up ubiquitous in a species very often, but it provably *does* happen. In fact, the Human DNA project has identified literally *thousands* of such fossilized "relics" of long-ago ancestral infections in the human DNA.
And several features of these DNA relics can be used to demonstrate common descent, including their *location*. The reason is that retroviruses aren't picky about where their DNA gets inserted into the host DNA. Even in an infection in a *single* individual, each infected cell has the retroviral DNA inserted into different locations than any other cell. Because the host DNA is so enormous (billions of basepairs in humans, for example), the odds of any retroviral insertion event matching the insertion location of any other insertion event are astronomically low. The only plausible mechanism by which two individuals could have retroviral DNA inserted into exactly the same location in their respective DNAs is if they inherited copies of that DNA from the same source -- a common ancestor.
Thus, shared endogenous retroviruses between, say, ape and man is almost irrefutable evidence that they descended from a common ancestor. *Unless* you want to suggest that they were created separately, and then a virus they were both susceptible to infected both a man and an ape in EXACTLY the same location in their DNAs (the odds of such a match by luck are literally on the order of 1,000,000,000,000,000,000 to 1...), *and* that the infections both happened in their gametocyte cells (combined odds on the order of 1,000,000 to 1) *and* that the one particular affected gametocyte is the one which produces the egg or sperm which is destined to produce an offspring (*HUGE* odds against), and *then* the resulting modified genome of the offspring becomes "fixed" in each respective population (1 out of population_size^squared)...
Then repeat that for *each* shared endogenous retrovirus (there are many) you'd like to claim was acquired independently and *not* from a shared ancestor...
Finally, you'd have to explain why, for say species A, B, and C, the pattern of shared same-location retroviruses is always *nested*, never *overlapped*. For example, all three will share some retroviruses, then A and B will both share several more, but if so then B *never* shares one with C that A doesn't also have (or at least remnants of).
In your "shared infection due to genetic similarities" suggestion, even leaving aside the near statistical impossibility of the infections leaving genetic "scars" in *exactly* the same locations in independent infections, one would expect to find cases of three species X, Y, and Z, where the degree of similarity was such that Y was "between" X and Z on some similarity scale, causing the same disease to befall X and Y but not Z, and another disease to affect Y and Z but not X. And yet, we don't find this in genetic markers. The markers are found in nested sequence, which is precisely what we would expect to see in cases of inheritance from common ancestry.
Here, for example, is an ancestry tree showing the pattern of shared same-location endogenous retroviruses of type HERV-K among primates:
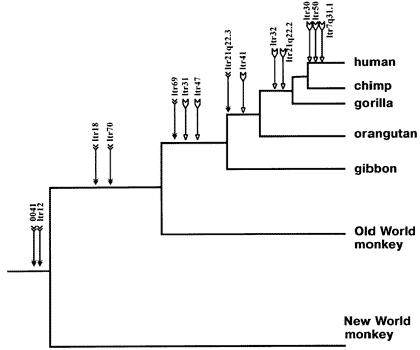
This is just a partial list for illustration purposes -- there are many more.
Each labeled arrow on the chart shows an ERV shared in common by all the branches to the right, and *not* the branches that are "left-and-down". This is the pattern that common descent would make. And common descent is the *only* plausible explanation for it. Furthermore, similar findings tie together larger mammal groups into successively larger "superfamilies" of creatures all descended from a common ancestor.
Any presumption of independent acquisition is literally astronomically unlikely. And "God chose to put broken relics of viral infections that never actually happened into our DNA and line them up only in patterns that would provide incredibly strong evidence of common descent which hadn't actually happened" just strains credulity (not to mention would raise troubling questions about God's motives for such a misleading act).
Once again, the evidence for common descent -- as opposed to any other conceivable alternative explanation -- is clear and overwhelming.
Wait, want more? Endogenous retroviruses are just *one* type of genetic "tag" that makes perfect sense evolutionary and *no* sense under any other scenario. In addition to ERV's, there are also similar arguments for the patterns across species of Protein functional redundancies, DNA coding redundancies, shared Processed pseudogenes, shared Transposons (including *several* independent varieties, such as SINEs and LINEs), shared redundant pseudogenes, etc. etc. Here, for example, is a small map of shared SINE events among various mammal groups:
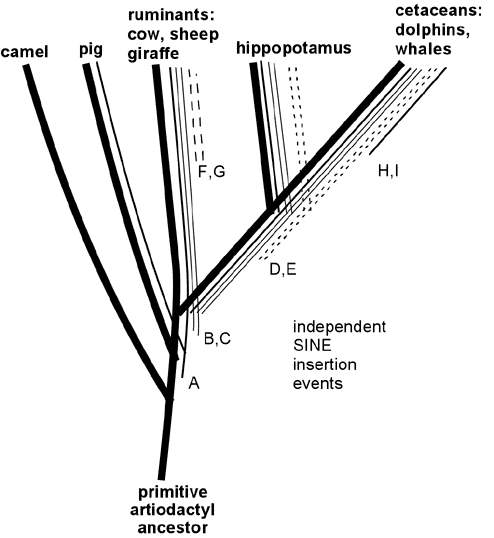
Like ERV's, any scenario which suggests that these shared DNA features were acquired separately strains the laws of probability beyond the breaking point, but they make perfect sense from an evolutionary common-descent scenario. In the above data, it is clear that the only logical conclusion is that, for example, the cetaceans, hippos, and ruminants shared a common ancestor, in which SINE events B and C entered its DNA and then was passed on to its descendants, yet this occurred after the point in time where an earlier common ancestor had given rise both to that species, and to the lineage which later became pigs.
And this pattern (giving the *same* results) is repeated over and over and over again when various kinds of molecular evidence from DNA is examined in detail.
The molecular evidence for evolution and common descent is overwhelming. The only alternative is for creationists to deny the obvious and say, "well maybe God decided to set up all DNA in *only* ways that were consistent with an evolutionary result even though He'd have a lot more options open to him, even including parts which by every measure are useless and exactly mimic copy errors, ancient infections, stutters, and other garbage inherited from nonexistent shared ancestors"...
Or how about:
Humans have 23 pairs of chromosomes ---chimps and gorillas have 24 pairs. How many pairs of chromosomes did the "common ancestor" have? Was it 23 or 24 pairs? How do you "evolve" missing or added chromosomes ---that would happen all at one time. The common ancestor had 24 chromosomes.
If you look at the gene sequences, you'll find that Chromosome 2 in humans is pretty much just 2 shorter chimpanzee chromosomes pasted end-to-end, with perhaps a slight bit of lost overlap:

(H=Human, C=Chimpanzee, G=Gorilla, O=Orangutan)
Somewhere along the line, after humans split off from the other great apes, or during the split itself, there was an accidental fusion of two chromosomes, end-to-end. Where there used to be 24 chromosomes, now there were 23, but containing the same total genes, so other than a "repackaging", the DNA "instructions" remained the same.
If a chimpanzee gives birth to a creature with 23 chromosomes, that offspring isn't going to be a well-formed chimpanzee able to survive well.
It is if the same genes are present, which they would be in the case of a chromosome fusion.
Evolve would imply the genetic material changes little by little --not some big loss of two chromosomes at once but I don't see how they'd go away gene by gene.
Tacking two chromosomes together end-to-end is not a "big loss" of genes, and it really is a "little by little" change in the total genetic code. It's just been "regrouped" a bit. Instead of coming in 24 "packages", it's now contained in 23, but the contents are the same.
So how, you might ask, would the chromosomes from the first 23-chromosome "fused" individual match up with the 24 chromosomes from its mate when it tried to produce offspring? Very well, thanks for asking. The "top half" of the new extra-long Chromosome 2 would adhere to the original chromosome (call it "2p") from which it was formed, and likewise for the "bottom half" which would adhere to the other original shorter chromosome (call it "2q"). In the picture above, imagine the two chimp chromosomes sliding over to "match up" against the human chromosome. The chimp chromosomes would end up butting ends with each other, or slightly overlapping in a "kink", but chromosomes have overcome worse mismatches (just consider the XY pair in every human male -- the X and the Y chromosome are *very* different in shape, length, and structure, but they still pair up).
In fact, the "rubbing ends" of the matched-up chimp chromosomes, adhering to the double-long human-type chromosome, would be more likely to become fused together themselves.
For studies in which recent chromosome fusions have been discovered and found not to cause infertility, see:
Chromosomal heterozygosity and fertility in house mice (Mus musculus domesticus) from Northern Italy. Hauffe HC, Searle JB Department of Zoology, University of Oxford, Oxford OX1 3PS, United Kingdom. hauffe@novanet.it An observed chromosome fusion: Hereditas 1998;129(2):177-80 A new centric fusion translocation in cattle: rob (13;19). Molteni L, De Giovanni-Macchi A, Succi G, Cremonesi F, Stacchezzini S, Di Meo GP, Iannuzzi L Institute of Animal Husbandry, Faculty of Agricultural Science, Milan, Italy.
J Reprod Fertil 1979 Nov;57(2):363-75 Cytogenetics and reproduction of sheep with multiple centric fusions (Robertsonian translocations). Bruere AN, Ellis PM
J Reprod Fertil Suppl 1975 Oct;(23):356-70 Cytogenetic studies of three equine hybrids. Chandley AC, Short RV, Allen WR.
In that last reference, the Przewalski horse, which has 33 chromosomes, and the domestic horse, with 32 chromosomes (due to a fusion), are able to mate and produce fertile offspring. Meanwhile, the question may be asked, how do we know that the human Chromosome 2 is actually the result of a chromsome fusion at/since a common ancestor, and not simply a matter of "different design"?
Well, if two chromsomes accidentally merged, there should be molecular remnants of the original chromosomal structures (while a chromosome designed from scratch would have no need for such leftover "train-wreck" pieces).
Ends of chromosomes have characteristic DNA base-pair sequences called "telomeres". And there are indeed remnants of telomeres at the point of presumed fusion on human Chromosome 2 (i.e., where the two ancestral ape chromosomes merged end-to-end). If I may crib from a web page:
Telomeres in humans have been shown to consist of head to tail repeats of the bases 5'TTAGGG running toward the end of the chromosome. Furthermore, there is a characteristic pattern of the base pairs in what is called the pre-telomeric region, the region just before the telomere. When the vicinity of chromosome 2 where the fusion is expected to occur (based on comparison to chimp chromosomes 2p and 2q) is examined, we see first sequences that are characteristic of the pre-telomeric region, then a section of telomeric sequences, and then another section of pre-telomeric sequences. Furthermore, in the telomeric section, it is observed that there is a point where instead of being arranged head to tail, the telomeric repeats suddenly reverse direction - becoming (CCCTAA)3' instead of 5'(TTAGGG), and the second pre-telomeric section is also the reverse of the first telomeric section. This pattern is precisely as predicted by a telomere to telomere fusion of the chimpanzee (ancestor) 2p and 2q chromosomes, and in precisely the expected location. Note that the CCCTAA sequence is the reversed complement of TTAGGG (C pairs with G, and T pairs with A).
Another piece of evidence is that if human Chromosome 2 had formed by chromosome fusion in an ancestor instead of being designed "as is", it should have evidence of 2 centromeres (the "pinched waist" in the picture above -- chromosomes have centromeres to aid in cell division). A "designed" chromosome would need only 1 centromere. An accidentally "merged" chromosome would show evidence of the 2 centromeres from the two chromosomes it merged from (one from each). And indeed, as documented in (Avarello R, Pedicini A, Caiulo A, Zuffardi O, Fraccaro M, Evidence for an ancestral alphoid domain on the long arm of human chromosome 2. Hum Genet 1992 May;89(2):247-9), the functional centromere found on human Chromosome 2 lines up with the centromere of the chimp 2p chromosome, while there are non-functional remnants of the chimp 2q centromere at the expected location on the human chromosome. As an aside, the next time some creationist claims that there is "no evidence" for common ancestry or evolution, keep in mind that the sort of detailed "detective story" discussed above is repeated literally COUNTLESS times in the ordinary pursuit of scientific research and examination of biological and other types of evidence. Common ancestry and evolution is confirmed in bit and little ways over and over and over again. It's not just something that a couple of whacky anti-religionists dream up out of thin air and promulgate for no reason, as the creationists would have you believe.
And:
[The poster known as Mr. LLLICHY wrote:] Here is that Vitamin C dataAfter discovering this same data on another thread along with more discussion than has appeared here (I've taken the liberty of pinging the participants of that discussion), I see what the "mystery" is supposed to be -- it's supposed be why did some sites have multiple mutations while (small) stretches of other sites had none? In other words, why do the mutations appear clustered?
(You know, it would really help if people explained their points and questions in more detail, instead of leaving people to guess what the poster was thinking...)
[LLLICHY wrote:] "U238" that decays thrice, pretty good trick when there is "U238" that does not decay at all in 50,000,000 years.
Actually, no site had mutations "thrice". Three different bases at a given site is only *two* mutations (one original base, plus two mutations from it to something else).
Here's the "mutation map" from the actual DNA data:
--1-12--1-1-1-1--------1112112--1---1-11-1--------1 ALL/n
No mutations ("-") in about half the sites, one mutation at several (17) sites, two mutations at three sites. The first thing to keep in mind that random processes tend to "cluster" more than people expect anyway. People expect "randomness" to "spread out" somewhat evenly, but instead it's usually more "clumped", for statistical reasons that would be a diversion to go into right now. So "that looks uneven" isn't always a good indication that something truly is non-random.
If you don't believe me on that, I wrote a program which made 23 mutations totally at random on a 51-site sequence, then repeated the process to see what different random outcomes would look like:
10 X$=STRING$(51,"-")
20 FOR I=1 TO 23
30 J%=INT(RND*51)+1
40 C$=MID$(X$,J%,1)
50 IF C$="-" THEN MID$(X$,J%,1)="1" ELSE MID$(X$,J%,1)=CHR$(ASC(C$)+1)
60 NEXT I
70 PRINT X$
80 GOTO 10
Yeah, it's BASIC, so sue me. Here's a typical screenful of the results: -21---1---2---111----2-----2-1121-------1---1--11-1
-1--1--21-11---1-1--1-1---1----1---21-11111---11---
3-11---3-----1-----11-2-1---1--1----3--2---1--1----
---1-1--22--1-1--2-2111--1-1111---1------1-------1-
---32----1-11-1-----1---2-231----1------1-----11--1
----2---21--1---4----1-------------11-1--111-11-211
11--1-1---1-----1--1------1----3111--1----111-2-1-2
1112---1-3-1----1-1-----1-1------121--111-------1-1
-111121--1----1----1-1-1-1-11-2---1-1-------1-111--
-----------11-1---11-11--------21----12211--1---131
--1-211-1-1----21--11-1-2----1--1----11---11-----11
12---1-13------------2---21-21---11-1-1-1--2-------
-----2-1---1-1----21--11-11-1---111-1--111-----2--1
-----1-----1-1-1-1---1-2----11-21-11--1-111---1-21-
---11--1-1-122-1-1-1--1-----2-1-1-1-------1-1---111
--2--11----2--1---12-2----1-1---1-1--1--12----1-1-1
-111-1-----1-1----------1-21111--1-2-11-11-1----11-
11-1--211-1221-----1--1-----11--1-2-1----------11--
-----1-12-11---2-1---11--1-2--1----11---111-1----11
11----1--12---12----1---31---1-11----2--1-11-1-----
---1--111-1--1-1-111----1-21----1-1-3---1------2--1
-2-11----1-1------1------2-1-1--111-111-1-1----1111
1--1--1-1---1-111111--2--1-1------112----2---11----
Notice how oddly "clustered" most of them look, including one run which left a 13-site stretch "absolutely untouched", contrary to intuition (while having *4* mutations at a single site!) Frankly, I don't see anything in the real-life DNA mutation map which looks any different from these truly random runs. Random events tend to cluster more than people expect. That solves the "mystery" right there.
Also, there may be a selection factor -- the GLO gene is a *lot* bigger than this. One has to wonder if this small 51-bp section was presented just because it was the one that looked "least random". That would be a no-no, since one can always hand-select the most deviant subset out of larger sample in order to artificially skew the picture.
However, since there are some interesting evolutionary observations to be made, let's look at that DNA data again, slightly rearranged:
TAC CCC GTG GAG GTG CGC TTC ACT CGG GCG GAC GAC ATC CTG CTG AGC CCC PIG
TAC CCC GTG GAG GTA CGC TTC ACT CGC GGG GAC GAC ATC CTG CTG AGC CCC BOS
TAC CCC GTA GAG GTG CGC TTC ACC CGA GGC GAT GAC ATT CTG CTG AGC CCC RAT
TAC CCC GTG GAG GTG CGC TTC ACC CGA GGT GAT GAC ATC CTG CTG AGC CCG MOUSE
TAC CCT GTG GGG GTG CGC TTC ACC CGG GGG GAC GAC ATC CTG CTG AGC CCC GUIN PIG
TAC CTG GTG GGG GTA CGC TTC ACC TGG AG* GAT GAC ATC CTA CTG AGC CCC HUMAN
TAC CTG GTG GGG CTA CGC TTC ACC TGG AG* GAT GAC ATC CTA CTG AGC CCC CHIMPANZEE
TAC CCG GTG GGG GTG CGC TTC ACC CAG AG* GAT GAC GTC CTA CTG AGC CCC ORANGUTAN
TAA CCG GTG GGG GTG CGC TTC ACC CAA GG* GAT GAC ATC ATA CTG AGC CCC MACAQUE
Here I've put spaces between codons, and clustered the closely-related species together: pig/cow as ungulates, rat/mouse for their obvious relationship, guinea pig right below them but separated because of the pseudogene nature of its GLO gene, then primates all in a group, with man's closest relative, the chimp, immediately below him, followed by the more distant orangutan, and the even more distant macaque. Also note that the top four have "working" GLO genes, and the bottom five have "broken" GLO pseudogenes. First, let's consider just the four species with working GLO genes. Evolution predicts that even over large periods of time, these genes will be "highly conserved", with natural selection weeding out mutations that could "break" the gene. Note that the mutations will still have occurred in individuals of the population, but natural selection will "discourage" that mutation from spreading into the general population.
And before we go any further, let's talk about the "universal genetic code". In all mammals (indeed, in almost all living organisms), each triplet of DNA sites cause a particular amino acid to be formed. The mapping of triplets (called "codons") to amino acids is as follows:
|
|
|
Second Position of Codon |
|
|
|
|
|
T |
C |
A |
G |
|
|
F
i
r
s
t
P
o
s
i
t
i
o
n |
T |
| TTT |
Phe |
[F] |
| TTC |
Phe |
[F] |
| TTA |
Leu |
[L] |
| TTG |
Leu |
[L] |
|
| TCT |
Ser |
[S] |
| TCC |
Ser |
[S] |
| TCA |
Ser |
[S] |
| TCG |
Ser |
[S] |
|
| TAT |
Tyr |
[Y] |
| TAC |
Tyr |
[Y] |
| TAA |
Ter |
[end] |
| TAG |
Ter |
[end] |
|
| TGT |
Cys |
[C] |
| TGC |
Cys |
[C] |
| TGA |
Ter |
[end] |
| TGG |
Trp |
[W] |
|
|
T
h
i
r
d
P
o
s
i
t
i
o
n |
| C |
| CTT |
Leu |
[L] |
| CTC |
Leu |
[L] |
| CTA |
Leu |
[L] |
| CTG |
Leu |
[L] |
|
| CCT |
Pro |
[P] |
| CCC |
Pro |
[P] |
| CCA |
Pro |
[P] |
| CCG |
Pro |
[P] |
|
| CAT |
His |
[H] |
| CAC |
His |
[H] |
| CAA |
Gln |
[Q] |
| CAG |
Gln |
[Q] |
|
| CGT |
Arg |
[R] |
| CGC |
Arg |
[R] |
| CGA |
Arg |
[R] |
| CGG |
Arg |
[R] |
|
|
| A |
| ATT |
Ile |
[I] |
| ATC |
Ile |
[I] |
| ATA |
Ile |
[I] |
| ATG |
Met |
[M] |
|
| ACT |
Thr |
[T] |
| ACC |
Thr |
[T] |
| ACA |
Thr |
[T] |
| ACG |
Thr |
[T] |
|
| AAT |
Asn |
[N] |
| AAC |
Asn |
[N] |
| AAA |
Lys |
[K] |
| AAG |
Lys |
[K] |
|
| AGT |
Ser |
[S] |
| AGC |
Ser |
[S] |
| AGA |
Arg |
[R] |
| AGG |
Arg |
[R] |
|
|
| G |
| GTT |
Val |
[V] |
| GTC |
Val |
[V] |
| GTA |
Val |
[V] |
| GTG |
Val |
[V] |
|
| GCT |
Ala |
[A] |
| GCC |
Ala |
[A] |
| GCA |
Ala |
[A] |
| GCG |
Ala |
[A] |
|
| GAT |
Asp |
[D] |
| GAC |
Asp |
[D] |
| GAA |
Glu |
[E] |
| GAG |
Glu |
[E] |
|
| GGT |
Gly |
[G] |
| GGC |
Gly |
[G] |
| GGA |
Gly |
[G] |
| GGG |
Gly |
[G] |
|
|
(The above table imported from http://psyche.uthct.edu/shaun/SBlack/geneticd.html, which also has a nice introduction to the genetic code.)
Another version of the same table with nifty Java features and DNA database lookups can be found here.
The thing which is most relevant to the following discussion is the fact that most of the genetic codes are "redundant" -- more than one codon (triplet) encodes to exactly the same amino acid. This means that even in genes which are required for the organism, certain basepair mutations make absolutely no difference if the change is from one codon which maps into amino acid X to another codon which still maps into amino acid X. (This fact allows certain kinds of evolutionary "tracers" to be "read" from the DNA, as described here).
Now back to our DNA data. The redundancy in the genetic code means that some basepair sites will have more "degrees of freedom" than others (i.e., ways in which they can mutate without disrupting the gene's biological function in any way). Let's look at the four species with working GLO genes again:
TAC CCC GTG GAG GTG CGC TTC ACT CGG GCG GAC GAC ATC CTG CTG AGC CCC PIG
TAC CCC GTG GAG GTA CGC TTC ACT CGC GGG GAC GAC ATC CTG CTG AGC CCC BOS
TAC CCC GTA GAG GTG CGC TTC ACC CGA GGC GAT GAC ATT CTG CTG AGC CCC RAT
TAC CCC GTG GAG GTG CGC TTC ACC CGA GGT GAT GAC ATC CTG CTG AGC CCG MOUSE
T T T A T A T T T A T C C T T T T T T T T
A A A A A C A A A A A
G C G G G G G C C C
--- --- --1 --- --1 --- --- --1 --2 -12 --1 --- --1 --- --- --- --1
Under each site of the mouse DNA, I've listed the "alternative" bases which could be be substituted for the mouse base at that site WITHOUT ALTERING THE GENE'S FUNCTION (because of genetic code redundancy). And under that I show the "mutation map" of just those four species. Note that most of the "alternative" bases are in the third base of each codon, *and* that this is where all but one of the mutations have appeared. This is because these were the sites which were "free" to mutate in the way they did, because the mutation was genetically neutral. That doesn't mean that the first and second sites of each codon were immune from mutation, it's just that when mutations did occur at those sites, natural selection weeded them out quickly because they most likely "broke" the GLO gene for the individuals which received that mutuation. What we see above is the results after natural selection has already "filtered" the undesirable mutations and left the ones which "do no harm".
Additionally, the two sites which have mutated twice (i.e. have a "2" in the mutation map) are ones which had more "allowable" mutations. Also note that the sites which had the fewest allowable alternatives (only one alternate letter allowed) didn't have any mutations fix at those sites, which is unsurprising since a "safe" mutation would be less likely to occur there versus a site that "allowed" two or three alternatives.
All this is as predicted by evolutionary theory, you'll note.
It also explains the one anomoly of the original mutation map, which is that the mutation counts do tend to be higher at the third base of a codon.
However... What about the one exception? The pig DNA has had one mutation at a site which does not encode to exactly the same amino acid (which is the case for *all* the other ones). In the pig DNA, the GGG codon (mapping to Glycine) has changed to a GCG codon (mapping to Alanine). What's up with that? Well, one of two things. First and most likely, just as base values in codons have a built-in redundancy, so do the amino acids which make up the proteins which result from the DNA templates. In other words, certain amino acids can be substituted for other ones at some sites in given proteins without making any functional difference. (This "protein functional redundancy" also has implications for "evolutionary tracer" analysis, see here.) That may well be the case for Alanine versus Glycine in the GLO protein, but I'm not enough of a biochemist to be able to say. The other option is that it *does* make some difference in the function of the pig GLO protein, but not enough to "break" the vitamin-C synthesis (as proven by the fact that pigs *can* synthesize vitamin C). So one way or another, it's not a deal-breaker even though pig GLO will not be 100% identical to cow/mouse/rat GLO. It's yet another "allowable" mutation.
More interesting evolutionary observations: The number of mutational differences between pig/cow is 3, the number between mouse/rat is 4, and the difference between rat/cow is 7 -- all roughly as one would expect from the evolutionary relatedness of these animals (cows/pigs and rats/mice are each closer to each other than the rodents are to the ungulates).
Now let's take a close look at the guinea pig:
TAC CCT GTG GGG GTG CGC TTC ACC CGG GGG GAC GAC ATC CTG CTG AGC CCC GUIN PIG
--- --1 --- -1- --- --- --- --- --1 --1 --1 --- --- --- --- --- ---
The "mutation map" under the guinea pig DNA is compared to the mouse DNA. Fascinating: Note that four of the five mutations are in the third base of a codon, *and* are of the type "allowed" by the genetic code redundancy. This indicates strongly that most of the evolutionary divergence between guinea pigs and mice likely occurred while the guinea pig's ancestors still had a working GLO gene. This is the sort of prediction implied by the evolutionary theory which could be cross-checked by further research of various types, and if verified, would be yet further confirmation that evolutionary theory is likely correct. So far, evolutionary theory has been subjected to literally countless tests like this, large and small, and the vast majority of results have confirmed the evolutionary prediction. This track record is hard to explain if evolution is an invalid theory, as some assert... Finally, let's look over the primate DNA and mutation map (relative to each other):
TAC CTG GTG GGG GTA CGC TTC ACC TGG AG* GAT GAC ATC CTA CTG AGC CCC HUMAN
TAC CTG GTG GGG CTA CGC TTC ACC TGG AG* GAT GAC ATC CTA CTG AGC CCC CHIMPANZEE
TAC CCG GTG GGG GTG CGC TTC ACC CAG AG* GAT GAC GTC CTA CTG AGC CCC ORANGUTAN
TAA CCG GTG GGG GTG CGC TTC ACC CAA GG* GAT GAC ATC ATA CTG AGC CCC MACAQUE
--1 -1- --- --- 1-1 --- --- --- 111 1-- --- --- 1-- 1-- --- --- ---
Evolutionary theory predicts that because the GLO gene is "broken" in primates (i.e. is a pseudogene), mutations in it are highly likely to be neutral (i.e., make no difference, since it can't get much more broken), and thus mutations are just as likely to accumulate at any site as any other. Is that what we see? Yup. There's no obvious pattern to the mutations between primates in the above mutation map, and unlike the pig/cow/mouse/rat mutation map, the mutations aren't predominantly at the "safer" third base of a codon, nor of a type that would be "safe". In fact, one base has vanished entirely, but no biggie, the gene's already broken. Also, although primates share a more recent common ancestor than cows/pigs/mice/rats, note that they've already racked up almost as many relative mutations as the cow/pig/mouse/rat DNA. This too is just as evolutionary theory predicts, because many mutations in a functional gene (GLO in this case) will be "non-safe" and weeded out by natural selection, making for a slower mutation fixation rate overall than in a pseudogene (as GLO is in primates) where natural selection doesn't "care" about the vast majority of mutations since *most* are neutral. So pseudogenes accumulate mutations faster than functional genes (even though rate of mutation *occurence* in both are likely the same).
Finally, note that there are ZERO mutational differences between the human DNA and the chimpanzee DNA, our nearest living relative.
I also see some interesting implications in the DNA sequences concerning which specific mutation fixed during what branch of the common-descent evolutionary tree for all the species represented, but reconstructing that would not only take another couple hours, at least, but would be a major bear to code in HTML, since I'd have to draw trees with annotations on the nodes... Bleugh.
In any case, I hope I've clarified some of the methods by which biologists find countless confirmations of evolution in DNA data. This is just a "baby" example, and to be more statistically valid would have to be done over much vaster sections of DNA sequences, but my intent was to demonstrate some of the concepts.
And if such a small amount of DNA as this can make small confirmations of evolutionary predictions, imagine the amount of confirmation from billion-basepair DNA data from each species compared across thousands of species... The amount of confirmatory discoveries for evolution from DNA analysis has already been vast, and promises to only grow in the future. For an overview of some of the different lines of evidence being studied, see The Journal of Molecular Evolution -- abstracts of all articles, current and back issues, can be browsed free online.
See also, for example (out of thousands):
Analysis of the human Alu Ye lineageHuman endogenous retrovirus HERV-K14 families: status, variants, evolution, and mobilization of other cellular sequences
Ancestral population sizes and species divergence times in the primate lineage on the basis of intron and BAC end sequences
Molecular and temporal characteristics of human retropseudogenes.
Evolution of RPS4Y
Inactivation of CMP-N-acetylneuraminic acid hydroxylase occurred prior to brain expansion during human evolution
Serine hydroxymethyltransferase pseudogene, SHMT-ps1: a unique genetic marker of the order primates
Structure and phylogenetic analysis of an endogenous retrovirus inserted into the human growth factor gene pleiotrophin
Molecular evolution of cytochrome c oxidase subunit IV: evidence for positive selection in simian primates
The ZNF75 zinc finger gene subfamily: isolation and mapping of the four members in humans and great apes
Insertions and duplications of mtDNA in the nuclear genomes of Old World monkeys and hominoids
Conservation of sequences between human and gorilla lineages: ADP-ribosyltransferase (NAD+) pseudogene 1 and neighboring retroposons
Fixation times of retroposons in the ribosomal DNA spacer of human and other primates
The gamma-globin genes and their flanking sequences in primates: findings with nucleotide sequences of capuchin monkey and tarsier
The emergence of new DNA repeats and the divergence of primates
Genetic diversity at class II DRB loci of the primate MHC
Nucleotide sequences of immunoglobulin-epsilon pseudogenes in man and apes and their phylogenetic relationships
Structure and evolution of human and African ape rDNA pseudogenes
How many more would you like?
To: frgoff; thomaswest
[And the existence of human fetal tails actually supports "macro" evolution.] Octogeny recapitulating Phylogeny has been discredited for probably over thirty years, now.
Try "ontogeny". You'll have a better chance of seeming like you know what you're talking about if you don't get the basic terminology wrong.
Furthermore, his point does not rest on recapitulation, so you either misunderstood his point, or are engaging in a red herring non sequitur.
Now would you care to engage his actual point, or not?
To: Aquinasfan
[Just require teachers to discuss evidence that refutes Charles Darwin's theory of evolution, ...] We can't have that because... uh, why not?
Because there isn't any?
Navigation: use the links below to view more comments.
first previous 1-20 ... 101-120, 121-140, 141-160 ... 201-213 next last
Disclaimer:
Opinions posted on Free Republic are those of the individual
posters and do not necessarily represent the opinion of Free Republic or its
management. All materials posted herein are protected by copyright law and the
exemption for fair use of copyrighted works.
FreeRepublic.com is powered by software copyright 2000-2008 John Robinson