That's a keeper...

Posted on 01/28/2005 4:28:41 PM PST by metacognative
Panicked Evolutionists: The Stephen Meyer Controversy
The theory of evolution is a tottering house of ideological cards that is more about cherished mythology than honest intellectual endeavor. Evolutionists treat their cherished theory like a fragile object of veneration and worship--and so it is. Panic is a sure sign of intellectual insecurity, and evolutionists have every reason to be insecure, for their theory is falling apart.
The latest evidence of this panic comes in a controversy that followed a highly specialized article published in an even more specialized scientific journal. Stephen C. Meyer, Director of the Discovery Institute's Center for Science and Culture, wrote an article accepted for publication in Proceedings of the Biological Society of Washington. The article, entitled "The Origin of Biological Information and the Higher Taxonomic Categories," was published after three independent judges deemed it worthy and ready for publication. The use of such judges is standard operating procedure among "peer-reviewed" academic journals, and is considered the gold standard for academic publication.
The readership for such a journal is incredibly small, and the Biological Society of Washington does not commonly come to the attention of the nation's journalists and the general public. Nevertheless, soon after Dr. Meyer's article appeared, the self-appointed protectors of Darwinism went into full apoplexy. Internet websites and scientific newsletters came alive with outrage and embarrassment, for Dr. Meyer's article suggested that evolution just might not be the best explanation for the development of life forms. The ensuing controversy was greater than might be expected if Dr. Meyer had argued that the world is flat or that hot is cold.
Eugenie C. Scott, Executive Director of the National Center for Science Education, told The Scientist that Dr. Meyer's article came to her attention when members of the Biological Society of Washington contacted her office. "Many members of the society were stunned about the article," she told The Scientist, and she described the article as "recycled material quite common in the intelligent design community." Dr. Scott, a well known and ardent defender of evolutionary theory, called Dr. Meyer's article "substandard science" and argued that the article should never have been published in any scientific journal.
Within days, the Biological Society of Washington, intimidated by the response of the evolutionary defenders, released a statement apologizing for the publication of the article. According to the Chronicle of Higher Education, the society's governing council claimed that the article "was published without the prior knowledge of the council." The statement went on to declare: "We have met and determined that all of us would have deemed this paper inappropriate for the pages of the Proceedings." The society's president, Roy W. McDiarmid, a scientist at the U.S. Geological Survey, blamed the article's publication on the journal's previous editor, Richard Sternberg, who now serves as a fellow at the National Center for Biotechnology Information at the National Institute of Health. "My conclusion on this," McDiarmid said, "was that it was a really bad judgment call on the editor's part."
What is it about Dr. Stephen Meyer's paper that has caused such an uproar? Meyer, who holds a Ph.D. from Cambridge University, argued in his paper that the contemporary form of evolutionary theory now dominant in the academy, known as "Neo-Darwinism," fails to account for the development of higher life forms and the complexity of living organisms. Pointing to what evolutionists identify as the "Cambrian explosion," Meyer argued that "the geologically sudden appearance of many new animal body plans" cannot be accounted for by Darwinian theory, "neo" or otherwise.
Accepting the scientific claim that the Cambrian explosion took place "about 530 million years ago," Meyer went on to explain that the "remarkable jump in the specified complexity or 'complex specified information' [CSI] of the biological world" cannot be explained by evolutionary theory.
The heart of Dr. Meyer's argument is found in this scientifically-loaded passage: "Neo-Darwinism seeks to explain the origin of new information, form, and structure as a result of selection acting on randomly arising variation at a very low level within the biological hierarchy, mainly, within the genetic text. Yet the major morphological innovations depend on a specificity of arrangement at a much higher level of the organizational hierarchy, a level that DNA alone does not determine. Yet if DNA is not wholly responsible for body plan morphogenesis, then DNA sequences can mutate indefinitely, without regard to realistic probabilistic limits, and still not produce a new body plan. Thus, the mechanism of natural selection acting on random mutations in DNA cannot in principle generate novel body plans, including those that first arose in the Cambrian explosion."
In simpler terms, the mechanism of natural selection, central to evolutionary theory, cannot possibly account for the development of so many varied and complex life forms simply by mutations in DNA. Rather, some conscious design--thus requiring a Designer--is necessary to explain the emergence of these life forms.
In the remainder of his paper, Meyer attacks the intellectual inadequacies of evolutionary theory and argues for what is now known as the "design Hypothesis." As he argued, "Conscious and rational agents have, as a part of their powers of purposive intelligence, the capacity to design information-rich parts and to organize those parts into functional information-rich systems and hierarchies." As he went on to assert, "We know of no other causal entity or process that has this capacity." In other words, the development of the multitude of higher life forms found on the planet can be explained only by the guidance of a rational agent--a Designer--whose plan is evident in the design.
Meyer's article was enough to cause hysteria in the evolutionists' camp. Knowing that their theory lacks intellectual credibility, the evolutionists respond by raising the volume, offering the equivalent of scientific shrieks and screams whenever their cherished theory is criticized--much less in one of their own cherished journals. As Dr. John West, Associate Director of the Discovery Institute explained, "Instead of addressing the paper's argument or inviting counterarguments or rebuttal, the society has resorted to affirming what amounts to a doctrinal statement in an effort to stifle scientific debate. They're trying to stop scientific discussion before it even starts."
When the Biological Society of Washington issued its embarrassing apology for publishing the paper, the organization pledged that arguments for Intelligent Design "will not be addressed in future issues of the Proceedings," regardless of whether the paper passes peer review.
From the perspective of panicked evolutionists, the Intelligent Design movement represents a formidable adversary and a constant irritant. The defenders of Intelligent Design are undermining evolutionary theory at multiple levels, and they refuse to go away. The panicked evolutionists respond with name-calling, labeling Intelligent Design proponents as "creationists," thereby hoping to prevent any scientific debate before it starts.
Intelligent Design is not tantamount to the biblical doctrine of creation. Theologically, Intelligent Design falls far short of requiring any affirmation of the doctrine of creation as revealed in the Bible. Nevertheless, it is a useful and important intellectual tool, and a scientific movement with great promise. The real significance of Intelligent Design theory and its related movement is the success with which it undermines the materialistic and naturalistic worldview central to the theory of evolution.
For the Christian believer, the Bible presents the compelling and authoritative case for God's creation of the cosmos. Specifically, the Bible provides us with the ultimate truth concerning human origins and the special creation of human beings as the creatures made in God's own image. Thus, though we believe in more than Intelligent Design, we certainly do not believe in less. We should celebrate the confusion and consternation now so evident among the evolutionists. Dr. Stephen Meyer's article--and the controversy it has spawned--has caught evolutionary scientists with their intellectual pants down.
_______________________________________
R. Albert Mohler, Jr
I'm aware there is no scientific explanation for the origin of life...or new organs.
In other words it is truth. We believe it is true because of our observations and witnesses. Whether it is true however, is separate from whether we accept it or whether it has been declared a law.
These evolving codes have intelligent designers.
Take off your blinders.
I knoe more about evolution than you do because I look at all sides. I follow the evidence ...not a 150 year old cult.
Not very well, apparently. After reading "Theory in Crisis" and "Icons of evolution". I began anew.
Oh, *there's* your problem. That's not "study" of evolution. That's like "studying" geopolitics by watching all of Michael Moore's movies.
Problems with Denton's arguments against evolution.
Problems with Wells's competence and honesty in his book.
If you're actually interested in *learning*, you'll read these links in their entirety with an open mind. If not, well, stick your head back in the sand.
It is a philosophical illusion...
Why, because a couple anti-evolutionist cranks can't get their facts right? I'm really not impressed.
maybe deliberate.
It's the conspiracy!!! Sooner or later, it seems most anti-evolutionists devolve into paranoia...
I would stand amidst your elephants unafraid.
Well, sure, as long as you keep your eyes tightly closed, you won't need be disturbed by any facts. A closed mind gathers no thought, and all that.
And their missing ancestors too!
What do you mean, "missing"?
(Most of the above text is from the link provided at the start of this post, and is the result of hard work by Kathleen Hunt, who deserves the credit. I've just extracted the relevant individual portions and assembled them into one direct fish-to-elephant sequence.) If you like that, here are a few hundred more.Fish to Amphibian transition: 1. Cheirolepis, (early Devonian, 400 million years ago) -- Primitive bony ray-finned fishes that gave rise to the vast majority of living fish. Heavy acanthodian-type scales, acanthodian-like skull, and big notocord.
2. Osteolepis (mid-Devonian, 390 million years ago) -- One of the earliest crossopterygian lobe-finned fishes, still sharing some characters with the lungfish (the other lobe-finned fishes). Had paired fins with a leg-like arrangement of major limb bones, capable of flexing at the "elbow", and had an early-amphibian-like skull and teeth.
3. Eusthenopteron, Sterropterygion (mid-late Devonian, 380 million years ago) -- Early rhipidistian lobe-finned fish roughly intermediate between early crossopterygian fish and the earliest amphibians. Skull very amphibian-like. Strong amphibian- like backbone. Fins very like early amphibian feet in the overall layout of the major bones, muscle attachments, and bone processes, with tetrapod-like tetrahedral humerus, and tetrapod-like elbow and knee joints. But there are no perceptible "toes", just a set of identical fin rays. Body & skull proportions rather fishlike.
4. Panderichthys, Elpistostege (mid-late Devonian, about 370 Mya) -- These "panderichthyids" are very tetrapod-like lobe-finned fish. Unlike Eusthenopteron, these fish actually look like tetrapods in overall proportions (flattened bodies, dorsally placed orbits, frontal bones! in the skull, straight tails, etc.) and have remarkably foot-like fins.
5. Obruchevichthys(middle Late Devonian, about 370 Mya -- Discovered in 1991 in Scotland, these are the earliest known tetrapod remains. The humerus is mostly tetrapod-like but retains some fish features. The discoverer, Ahlberg (1991), said: "It [the humerus] is more tetrapod-like than any fish humerus, but lacks the characteristic early tetrapod 'L-shape'...this seems to be a primitive, fish-like character....although the tibia clearly belongs to a leg, the humerus differs enough from the early tetrapod pattern to make it uncertain whether the appendage carried digits or a fin. At first sight the combination of two such extremities in the same animal seems highly unlikely on functional grounds. If, however, tetrapod limbs evolved for aquatic rather than terrestrial locomotion, as recently suggested, such a morphology might be perfectly workable."
6. Hynerpeton, Acanthostega, Ichthyostega (late Devonian, 360 Mya) -- A little later, the fin-to-foot transition was almost complete, and we have a set of early tetrapod fossils that clearly did have feet. The most complete are Ichthyostega, Acanthostega gunnari, and the newly described Hynerpeton bassetti (Daeschler et al., 1994). (There are also other genera known from more fragmentary fossils.) Hynerpeton is the earliest of these three genera (365 Ma), but is more advanced in some ways; the other two genera retained more fish- like characters longer than the Hynerpeton lineage did. Acanthostega still had internal gills, adding further support to the suggestion that unique tetrapod characters such as limbs with digits evolved first for use in water rather than for walking on land. Acanthostega also had a remarkably fish-like shoulder and forelimb. Ichthyostega was also very fishlike, retaining a fish-like finned tail, permanent lateral line system, and notochord. It turns out that Acanthostega's front foot had eight toes, and Ichthyostega's hind foot had seven toes, giving both feet the look of a short, stout flipper with many "toe rays" similar to fin rays. All you have to do to a lobe- fin to make it into a many-toed foot like this is curl it, wrapping the fin rays forward around the end of the limb. In fact, this is exactly how feet develop in larval amphibians, from a curled limb bud. Hynerpeton, in contrast, probably did not have internal gills and already had a well-developed shoulder girdle; it could elevate and retract its forelimb strongly, and it had strong muscles that attached the shoulder to the rest of the body (Daeschler et al., 1994).
7. Labyrinthodonts (eg Pholidogaster, Pteroplax) (late Dev./early Miss., 355 Mya) -- These larger amphibians still have some icthyostegid fish features, such as skull bone patterns, labyrinthine tooth dentine, presence & pattern of large palatal tusks, the fish skull hinge, pieces of gill structure between cheek & shoulder, and the vertebral structure. But they have lost several other fish features: the fin rays in the tail are gone, the vertebrae are stronger and interlocking, the nasal passage for air intake is well defined, etc.
Amphibian to Reptile transition: 8. Pholidogaster (Mississippian, about 330 Ma) -- A group of large labrinthodont amphibians, transitional between the early amphibians (the ichthyostegids, described above) and later amphibians such as rhachitomes and anthracosaurs.
9. Proterogyrinus (late Mississippian, 325 Mya) -- Classic labyrinthodont-amphibian skull and teeth, but with reptilian vertebrae, pelvis, humerus, and digits. Still has fish skull hinge. Amphibian ankle. 5-toed hand and a 2-3-4-5-3 (almost reptilian) phalangeal count.
10. Limnoscelis, Tseajaia (late Carboniferous, 300 Mya) -- Amphibians apparently derived from the early anthracosaurs, but with additional reptilian features: structure of braincase, reptilian jaw muscle, expanded neural arches.
11. Solenodonsaurus (mid-Pennsylvanian) -- An incomplete fossil, apparently between the anthracosaurs and the cotylosaurs. Loss of palatal fangs, loss of lateral line on head, etc. Still just a single sacral vertebra, though.
12. Hylonomus, Paleothyris (early Pennsylvanian) -- These are protorothyrids, very early cotylosaurs (primitive reptiles). They were quite little, lizard-sized animals with amphibian-like skulls (amphibian pineal opening, dermal bone, etc.), shoulder, pelvis, & limbs, and intermediate teeth and vertebrae. Rest of skeleton reptilian, with reptilian jaw muscle, no palatal fangs, and spool-shaped vertebral centra. Probably no eardrum yet.
13. Paleothyris (early Pennsylvanian) -- An early captorhinomorph reptile, with no temporal fenestrae at all.
14. Protoclepsydrops haplous (early Pennsylvanian) -- The earliest known synapsid reptile. Little temporal fenestra, with all surrounding bones intact. Had amphibian-type vertebrae with tiny neural processes. (reptiles had only just separated from the amphibians)
15. Clepsydrops (early Pennsylvanian) -- The second earliest known synapsid.
Reptile to Mammal transition: 16. Archaeothyris (early-mid Pennsylvanian) -- A slightly later ophiacodont. Small temporal fenestra, now with some reduced bones (supratemporal). Braincase still just loosely attached to skull. Slight hint of different tooth types. Still has some extremely primitive, amphibian/captorhinid features in the jaw, foot, and skull. Limbs, posture, etc. typically reptilian, though the ilium (major hip bone) was slightly enlarged.
17. Varanops (early Permian) -- Temporal fenestra further enlarged. Braincase floor shows first mammalian tendencies & first signs of stronger attachment to rest of skull (occiput more strongly attached). Lower jaw shows first changes in jaw musculature (slight coronoid eminence). Body narrower, deeper: vertebral column more strongly constructed. Ilium further enlarged, lower-limb musculature starts to change (prominent fourth trochanter on femur). This animal was more mobile and active. Too late to be a true ancestor, and must be a "cousin".
18. Haptodus (late Pennsylvanian) -- One of the first known sphenacodonts, showing the initiation of sphenacodont features while retaining many primitive features of the ophiacodonts. Occiput still more strongly attached to the braincase. Teeth become size-differentiated, with biggest teeth in canine region and fewer teeth overall. Stronger jaw muscles. Vertebrae parts & joints more mammalian. Neural spines on vertebrae longer. Hip strengthened by fusing to three sacral vertebrae instead of just two. Limbs very well developed.
19. Dimetrodon, Sphenacodon or a similar sphenacodont (late Pennsylvanian to early Permian, 270 Ma) -- More advanced pelycosaurs, clearly closely related to the first therapsids (next). Dimetrodon is almost definitely a "cousin" and not a direct ancestor, but as it is known from very complete fossils, it's a good model for sphenacodont anatomy. Medium-sized fenestra. Teeth further differentiated, with small incisors, two huge deep- rooted upper canines on each side, followed by smaller cheek teeth, all replaced continuously. Fully reptilian jaw hinge. Lower jaw bone made of multiple bones & with first signs of a bony prong later involved in the eardrum, but there was no eardrum yet, so these reptiles could only hear ground-borne vibrations (they did have a reptilian middle ear). Vertebrae had still longer neural spines (spectacularly so in Dimetrodon, which had a sail), and longer transverse spines for stronger locomotion muscles.
20. Biarmosuchia (late Permian) -- A therocephalian -- one of the earliest, most primitive therapsids. Several primitive, sphenacodontid features retained: jaw muscles inside the skull, platelike occiput, palatal teeth. New features: Temporal fenestra further enlarged, occupying virtually all of the cheek, with the supratemporal bone completely gone. Occipital plate slanted slightly backwards rather than forwards as in pelycosaurs, and attached still more strongly to the braincase. Upper jaw bone (maxillary) expanded to separate lacrymal from nasal bones, intermediate between early reptiles and later mammals. Still no secondary palate, but the vomer bones of the palate developed a backward extension below the palatine bones. This is the first step toward a secondary palate, and with exactly the same pattern seen in cynodonts. Canine teeth larger, dominating the dentition. Variable tooth replacement: some therocephalians (e.g Scylacosaurus) had just one canine, like mammals, and stopped replacing the canine after reaching adult size. Jaw hinge more mammalian in position and shape, jaw musculature stronger (especially the mammalian jaw muscle). The amphibian-like hinged upper jaw finally became immovable. Vertebrae still sphenacodontid-like. Radical alteration in the method of locomotion, with a much more mobile forelimb, more upright hindlimb, & more mammalian femur & pelvis. Primitive sphenacodontid humerus. The toes were approaching equal length, as in mammals, with #toe bones varying from reptilian to mammalian. The neck & tail vertebrae became distinctly different from trunk vertebrae. Probably had an eardrum in the lower jaw, by the jaw hinge.
21. Procynosuchus (latest Permian) -- The first known cynodont -- a famous group of very mammal-like therapsid reptiles, sometimes considered to be the first mammals. Probably arose from the therocephalians, judging from the distinctive secondary palate and numerous other skull characters. Enormous temporal fossae for very strong jaw muscles, formed by just one of the reptilian jaw muscles, which has now become the mammalian masseter. The large fossae is now bounded only by the thin zygomatic arch (cheekbone to you & me). Secondary palate now composed mainly of palatine bones (mammalian), rather than vomers and maxilla as in older forms; it's still only a partial bony palate (completed in life with soft tissue). Lower incisor teeth was reduced to four (per side), instead of the previous six (early mammals had three). Dentary now is 3/4 of lower jaw; the other bones are now a small complex near the jaw hinge. Jaw hinge still reptilian. Vertebral column starts to look mammalian: first two vertebrae modified for head movements, and lumbar vertebrae start to lose ribs, the first sign of functional division into thoracic and lumbar regions. Scapula beginning to change shape. Further enlargement of the ilium and reduction of the pubis in the hip. A diaphragm may have been present.
22. Dvinia [also "Permocynodon"] (latest Permian) -- Another early cynodont. First signs of teeth that are more than simple stabbing points -- cheek teeth develop a tiny cusp. The temporal fenestra increased still further. Various changes in the floor of the braincase; enlarged brain. The dentary bone was now the major bone of the lower jaw. The other jaw bones that had been present in early reptiles were reduced to a complex of smaller bones near the jaw hinge. Single occipital condyle splitting into two surfaces. The postcranial skeleton of Dvinia is virtually unknown and it is not therefore certain whether the typical features found at the next level had already evolved by this one. Metabolic rate was probably increased, at least approaching homeothermy.
23. Thrinaxodon (early Triassic) -- A more advanced "galesaurid" cynodont. Further development of several of the cynodont features seen already. Temporal fenestra still larger, larger jaw muscle attachments. Bony secondary palate almost complete. Functional division of teeth: incisors (four uppers and three lowers), canines, and then 7-9 cheek teeth with cusps for chewing. The cheek teeth were all alike, though (no premolars & molars), did not occlude together, were all single- rooted, and were replaced throughout life in alternate waves. Dentary still larger, with the little quadrate and articular bones were loosely attached. The stapes now touched the inner side of the quadrate. First sign of the mammalian jaw hinge, a ligamentous connection between the lower jaw and the squamosal bone of the skull. The occipital condyle is now two slightly separated surfaces, though not separated as far as the mammalian double condyles. Vertebral connections more mammalian, and lumbar ribs reduced. Scapula shows development of a new mammalian shoulder muscle. Ilium increased again, and all four legs fully upright, not sprawling. Tail short, as is necessary for agile quadrupedal locomotion. The whole locomotion was more agile. Number of toe bones is 2.3.4.4.3, intermediate between reptile number (2.3.4.5.4) and mammalian (2.3.3.3.3), and the "extra" toe bones were tiny. Nearly complete skeletons of these animals have been found curled up - a possible reaction to conserve heat, indicating possible endothermy? Adults and juveniles have been found together, possibly a sign of parental care. The specialization of the lumbar area (e.g. reduction of ribs) is indicative of the presence of a diaphragm, needed for higher O2 intake and homeothermy. NOTE on hearing: The eardrum had developed in the only place available for it -- the lower jaw, right near the jaw hinge, supported by a wide prong (reflected lamina) of the angular bone. These animals could now hear airborne sound, transmitted through the eardrum to two small lower jaw bones, the articular and the quadrate, which contacted the stapes in the skull, which contacted the cochlea. Rather a roundabout system and sensitive to low-frequency sound only, but better than no eardrum at all! Cynodonts developed quite loose quadrates and articulars that could vibrate freely for sound transmittal while still functioning as a jaw joint, strengthened by the mammalian jaw joint right next to it. All early mammals from the Lower Jurassic have this low-frequency ear and a double jaw joint. By the middle Jurassic, mammals lost the reptilian joint (though it still occurs briefly in embryos) and the two bones moved into the nearby middle ear, became smaller, and became much more sensitive to high-frequency sounds.
24. Cynognathus (early Triassic, 240 Ma; suspected to have existed even earlier) -- We're now at advanced cynodont level. Temporal fenestra larger. Teeth differentiating further; cheek teeth with cusps met in true occlusion for slicing up food, rate of replacement reduced, with mammalian-style tooth roots (though single roots). Dentary still larger, forming 90% of the muscle-bearing part of the lower jaw. TWO JAW JOINTS in place, mammalian and reptilian: A new bony jaw joint existed between the squamosal (skull) and the surangular bone (lower jaw), while the other jaw joint bones were reduced to a compound rod lying in a trough in the dentary, close to the middle ear. Ribs more mammalian. Scapula halfway to the mammalian condition. Limbs were held under body. There is possible evidence for fur in fossil pawprints.
25. Diademodon (early Triassic, 240 Ma; same strata as Cynognathus) -- Temporal fenestra larger still, for still stronger jaw muscles. True bony secondary palate formed exactly as in mammals, but didn't extend quite as far back. Turbinate bones possibly present in the nose (warm-blooded?). Dental changes continue: rate of tooth replacement had decreased, cheek teeth have better cusps & consistent wear facets (better occlusion). Lower jaw almost entirely dentary, with tiny articular at the hinge. Still a double jaw joint. Ribs shorten suddenly in lumbar region, probably improving diaphragm function & locomotion. Mammalian toe bones (2.3.3.3.3), with closely related species still showing variable numbers.
26. Probelesodon (mid-Triassic; South America) -- Fenestra very large, still separate from eyesocket (with postorbital bar). Secondary palate longer, but still not complete. Teeth double-rooted, as in mammals. Nares separated. Second jaw joint stronger. Lumbar ribs totally lost; thoracic ribs more mammalian, vertebral connections very mammalian. Hip & femur more mammalian.
27. Probainognathus (mid-Triassic, 239-235 Ma, Argentina) -- Larger brain with various skull changes: pineal foramen ("third eye") closes, fusion of some skull plates. Cheekbone slender, low down on the side of the eye socket. Postorbital bar still there. Additional cusps on cheek teeth. Still two jaw joints. Still had cervical ribs & lumbar ribs, but they were very short. Reptilian "costal plates" on thoracic ribs mostly lost. Mammalian #toe bones.
28. Pachygenelus, Diarthrognathus (earliest Jurassic, 209 Ma) -- These are trithelodontids. Inflation of nasal cavity, establishment of Eustachian tubes between ear and pharynx, loss of postorbital bar. Alternate replacement of mostly single- rooted teeth. This group also began to develop double tooth roots -- in Pachygenelus the single root of the cheek teeth begins to split in two at the base. Pachygenelus also has mammalian tooth enamel, and mammalian tooth occlusion. Double jaw joint, with the second joint now a dentary-squamosal (instead of surangular), fully mammalian. Incipient dentary condyle. Reptilian jaw joint still present but functioning almost entirely in hearing; postdentary bones further reduced to tiny rod of bones in jaw near middle ear; probably could hear high frequencies now. More mammalian neck vertebrae for a flexible neck. Hip more mammalian, with a very mammalian iliac blade & femur. Highly mobile, mammalian-style shoulder. Probably had coupled locomotion & breathing.
29. Sinoconodon (early Jurassic, 208 Ma) -- The next known very ancient proto-mammal. Eyesocket fully mammalian now (closed medial wall). Hindbrain expanded. Permanent cheekteeth, like mammals, but the other teeth were still replaced several times. Mammalian jaw joint stronger, with large dentary condyle fitting into a distinct fossa on the squamosal. This final refinement of the joint automatically makes this animal a true "mammal". Reptilian jaw joint still present, though tiny.
Proto-mammal to Placental Mammal transition: 30. Kuehneotherium (early Jurassic, about 205 Ma) -- A slightly later proto-mammal, sometimes considered the first known pantothere (primitive placental-type mammal). Teeth and skull like a placental mammal. The three major cusps on the upper & lower molars were rotated to form interlocking shearing triangles as in the more advanced placental mammals & marsupials. Still has a double jaw joint, though.
31. Eozostrodon, Morganucodon, Haldanodon (early Jurassic, ~205 Ma) -- A group of early proto-mammals called "morganucodonts". The restructuring of the secondary palate and the floor of the braincase had continued, and was now very mammalian. Truly mammalian teeth: the cheek teeth were finally differentiated into simple premolars and more complex molars, and teeth were replaced only once. Triangular- cusped molars. Reversal of the previous trend toward reduced incisors, with lower incisors increasing to four. Tiny remnant of the reptilian jaw joint. Once thought to be ancestral to monotremes only, but now thought to be ancestral to all three groups of modern mammals -- monotremes, marsupials, and placentals.
32. Peramus (late Jurassic, about 155 Ma) -- A "eupantothere" (more advanced placental-type mammal). The closest known relative of the placentals & marsupials. Triconodont molar has with more defined cusps. This fossil is known only from teeth, but judging from closely related eupantotheres (e.g. Amphitherium) it had finally lost the reptilian jaw joint, attaing a fully mammalian three-boned middle ear with excellent high-frequency hearing. Has only 8 cheek teeth, less than other eupantotheres and close to the 7 of the first placental mammals. Also has a large talonid on its "tribosphenic" molars, almost as large as that of the first placentals -- the first development of grinding capability.
33. Endotherium (very latest Jurassic, 147 Ma) -- An advanced eupantothere. Fully tribosphenic molars with a well- developed talonid. Known only from one specimen. From Asia; recent fossil finds in Asia suggest that the tribosphenic molar evolved there.
34. Vincelestes neuquenianus (early Cretaceous, 135 Ma) -- A probably-placental mammal with some marsupial traits, known from some nice skulls. Placental-type braincase and coiled cochlea. Its intracranial arteries & veins ran in a composite monotreme/placental pattern derived from homologous extracranial vessels in the cynodonts. (Rougier et al., 1992)
35. Kennalestes and Asioryctes (late Cretaceous, Mongolia) -- Small, slender animals; eyesocket open behind; simple ring to support eardrum; primitive placental-type brain with large olfactory bulbs; basic primitive tribosphenic tooth pattern. Canine now double rooted. Still just a trace of a non-dentary bone, the coronoid, on the otherwise all-dentary jaw. "Could have given rise to nearly all subsequent placentals." says Carroll (1988).
Placental mammal to elephant transition: 36. Protungulatum (latest Cretaceous) -- Transitional between earliest placental mammals and the condylarths (primitive, small hoofed animals). These early, simple insectivore- like small mammals had one new development: their cheek teeth had grinding surfaces instead of simple, pointed cusps. They were the first mammal herbivores. All their other features are generalized and primitive -- simple plantigrade five-toed clawed feet, all teeth present (3:1:4:3) with no gaps, all limb bones present and unfused, pointy-faced, narrow small brain, eyesocket not closed.
37. Minchenella or a similar condylarth (late Paleocene) -- Known only from lower jaws. Has a distinctive broadened shelf on the third molar.
38. Phenacolophus (late Paleocene or early Eocene) -- An early embrithopod (very early, slightly elephant-like condylarths), thought to be the stem-group of all elephants.
39. Pilgrimella (early Eocene) -- An anthracobunid (early proto-elephant condylarth), with massive molar cusps aligned in two transverse ridges.
40. Unnamed species of proto-elephant (early Eocene) -- Discovered recently in Algeria. Had slightly enlarged upper incisors (the beginnings of tusks), and various tooth reductions. Still had "normal" molars instead of the strange multi-layered molars of modern elephants. Had the high forehead and pneumatized skull bones of later elephants, and was clearly a heavy-boned, slow animal. Only one meter tall.
41. Moeritherium, Numidotherium, Barytherium (early-mid Eocene) -- A group of three similar very early elephants. It is unclear which of the three came first. Pig-sized with stout legs, broad spreading feet and flat hooves. Elephantish face with the eye set far forward & a very deep jaw. Second incisors enlarged into short tusks, in upper and lower jaws; little first incisors still present; loss of some teeth. No trunk.
42. Paleomastodon, Phiomia (early Oligocene) -- The first "mastodonts", a medium-sized animals with a trunk, long lower jaws, and short upper and lower tusks. Lost first incisors and canines. Molars still have heavy rounded cusps, with enamel bands becoming irregular. Phiomia was up to eight feet tall.
43. Gomphotherium (early Miocene) -- Basically a large edition of Phiomia, with tooth enamel bands becoming very irregular. Two long rows cusps on teeth became cross- crests when worn down. Gave rise to several families of elephant- relatives that spread all over the world. From here on the elephant lineages are known to the species level.
44a. The mastodon lineage split off here, becoming more adapted to a forest browser niche, and going through Miomastodon (Miocene) and Pliomastodon (Pliocene), to Mastodon (or "Mammut", Pleistocene).
44b. Meanwhile, the elephant lineage became still larger, adapting to a savannah/steppe grazer niche:
45. Stegotetrabelodon (late Miocene) -- One of the first of the "true" elephants, but still had two long rows of cross-crests, functional premolars, and lower tusks. Other early Miocene genera show compression of the molar cusps into plates (a modern feature ), with exactly as many plates as there were cusps. Molars start erupting from front to back, actually moving forward in the jaw throughout life.
46. Primelephas (latest Miocene) -- Short lower jaw makes it look like an elephant now. Reduction & loss of premolars. Very numerous plates on the molars, now; we're now at the modern elephants' bizarre system of one enormous multi-layered molar being functional at a time, moving forward in the jaw.
47. Primelephas gomphotheroides (mid-Pliocene) -- A later species that split into three lineages, Loxodonta, Elephas, and Mammuthus:
The Pleistocene record for elephants is very good. In general, after the earliest forms of the three modern genera appeared, they show very smooth, continuous evolution with almost half of the speciation events preserved in fossils. For instance, Carroll (1988) says: "Within the genus Elephas, species demonstrate continuous change over a period of 4.5 million years. ...the elephants provide excellent evidence of significant morphological change within species, through species within genera, and through genera within a family...."
- Loxodonta adaurora (5 Ma). Gave rise to the modern African elephant Loxodonta africana about 3.5 Ma.
- Elephas ekorensis (5 Ma), an early Asian elephant with rather primitive molars, clearly derived directly from P. gomphotheroides. Led directly to:
- Elephas recki, which sent off one side branch, E. hydrusicus, at 3.8 Ma, and then continued changing on its own until it became E. iolensis.
- Elephas maximus, the modern Asian elephant, clearly derived from
- E. hysudricus. Strikingly similar to young E. hysudricus animals. Possibly a case of neoteny (in which "new" traits are simply juvenile features retained into adulthood).
- Mammuthus meridionalis, clearly derived from P. gomphotheroides. Spread around the northern hemisphere. In Europe, led to M. armeniacus/trogontherii, and then to M. primigenius. In North America, led to M. imperator and then M. columbi.
Species-species transitions among the elephants:
- Maglio (1973) studied Pleistocene elephants closely. Overall, Maglio showed that at least 7 of the 17 Quaternary elephant species arose through smooth anagenesis transitions from their ancestors. For example, he said that Elephas recki "can be traced through a progressive series of stages...These stages pass almost imperceptibly into each other....In the late Pleistocene a more progressive elephant appears which I retain as a distinct species, E. iolensis, only as a matter of convenience. Although as a group, material referred to E. iolensis is distinct from that of E. recki, some intermediate specimens are known, and E. iolensis seems to represent a very progressive, terminal stage in the E. recki specific lineage."
- Maglio also documented very smooth transitions between three Eurasian mammoth species: Mammuthus meridionalis --> M. armeniacus (or M. trogontherii) --> M. primigenius.
- Lister (1993) reanalyzed mammoth teeth and confirmed Maglio's scheme of gradual evolution in European mammoths, and found evidence for gradual transitions in the North American mammoths too.
Similar fossil sequences can be listed for the majority of other major-group transitions.
(Did I hear a creationist in the back row say something about "no transitional fossils?")
Also note that the changes between any two sequential transitionals are small enough that most creationists would write them off as only "microevolution" -- and yet those 50-or-so "microevolutionary" steps turn a fish into an elephant, which even the most stubborn creationist would have to concede is "macroevolution".
Post #185, son. You couldn't *possibly* have missed it, so why are you faking amnesia now? That's not very honest of you.
ROFL!! If they "refuse to debate", then how did they "always lose"? Do you even think things through before you write them?
In any case, science is not decided by stage-show "debates". This ain't politics. And "debates" on stage are crappy venues for presenting the kind of volumes of evidence and analysis by which science is actually done. That's why scientific debates (*real* ones, not the "play-to-the-audience" kind Gish et al like to do as publicity stunts) are done in journals and forums.
Are you really this ignorant of the process? Do you really think that scientific theories can be properly vetted in a 30-minute stage show? If so... I'm wasting my time. And you've wasted your own.
I went back to 185. It was as useful as your elephants.
You're a fanatic if you think any of your hurls refute Wells or Denton.
No, actually, they don't. Please correct your ignorance. "These evolving codes" begin totally at random, and then randomly explore problem space while the alleged "intelligent designers" of the "codes" are out eating dinner.
Furthermore, the "evolving codes" can solve problems that even the *programmers* don't know how to solve. So where and how exactly did the "intelligent designers" put in the "intelligence" to solve the problem when THEY DIDN'T HAVE THAT INTELLIGENCE THEMSELVES?
Ponder that one for a while.
Look, I know what you're trying to say, but you're quite simply way off base. Tell me: If I use a telescope to examine a distant moon of Jupiter, does that mean that it's impossible for the moon to circle its planet because I used "intelligent design" to make the telescope I'm using gather data? If I build a wooden ramp to study how objects roll downhill, does that mean that no objects can roll down hills (or no hills can exist) without the kind of "intelligent design" I put into setting up the ramp for the experiment? This is *exactly* the kind of nonsense that anti-evolutionists spew when they try to hand-wave away the results of evolutionary algorithms by whining about how someone used their "intelligent design" to set up the experiment and monitor it as it runs (and runs BY ITSELF, I repeat again).
It's hogwash. I've *DONE* genetic algorithms. I *know* that I constructed the "experiment" in a way that precludes me "putting" the solution into the system, in just the same way that any good experimenter knows how to isolate the thing he's testing from his own interference. And they've usually come up with results that surprised even *me*, because not even *I* had the "intelligence" which the evolutionary processes CAME UP WITH ON THEIR OWN.
Honestly, you *don't* know how these things work, because you're frantically looking for *excuses* for why you think you shouldn't have to actually learn about them, instead of just *reading* the material and learning *from* it. It's pathetic, frankly, but I've come to expect it from anti-evolutionists. (That's how they *stay* anti-evolutionists, by hiding from the actual topic and the evidence for it.)
Take off your blinders.
Oh, the irony...
That's a keeper...
8-) And the peppered moth fiasco was "corner cutting" too?
CONGRATULATIONS!

Remember just a few minutes ago when I warned you thusly?
Well, you chose to ignore my advice, and so you've just made a fool of yourself, because there's NOTHING WRONG with the peppered moth example, and the link I gave you explains at length why Wells has his head up his butt when he claims there is. Here's what you neglected to read before you shot your mouth off (DESPITE my clear warning to you and link to the information):Friendly warning: If you've made the mistake of reading Wells "Icons of Evolution" anti-evolution book -- and *believing* it -- you need to read these before you attempt to respond: Icons of Evolution FAQs . They document just how wildly off-base (and how much of a *documented FRAUD*) Wells himself is in his bizarre accusations. Don't say I didn't warn you.
Chapter 7: Peppered Moths
So many things are wrong with Wells's treatment of peppered moths (Biston betularia) that it is hard to list them all; but I will try. The authoritative reference on this topic is Michael Majerus' 1998 book Melanism: Evolution in Action. This book includes two long chapters on Biston. The first chapter, "The peppered moth story," recounts the basic story of melanism in Biston, and relates how this story was pieced together by Kettlewell and others. The second chapter, "The peppered moth story dissected," gives a thorough critical review of the basic story, considering aspects and details of the basic story in the light of research (by Majerus and others) post-dating Kettlewell.
Crucially, however, Majerus clearly and explicitly concludes that, in his view, Kettlewell got things basically correct. At the beginning of his second peppered moth chapter, Majerus writes,
First, it is important to emphasize that, in my view, the huge wealth of additional data obtained since Kettlewell's initial predation papers (Kettlewell 1955a, 1956) does not undermine the basic qualitative deductions from that work. Differential bird predation of the typica and carbonaria forms, in habitats affected by industrial pollution to different degrees, is the primary influence of the evolution of melanism in the peppered moth (Majerus, 1998, p. 116).
Majerus is so clear on this point that one suspects that he was anticipating that his critique would be misinterpreted by non-peppered moth researchers. It seems that there is a "too good to be true" quality about the peppered moth story that leads people to interpret any hint of criticism as a sign that the whole basic story is crashing down. Scientists are by no means immune to this tendency, and indeed they may be more prone to it given the regularity with which popular ideas have been overturned throughout the history of science. The press has an even greater tendency towards snap judgements and oversimplifications when it comes to scientific discussions. Antievolutionists, on the other hand, have always been stuck muttering "it's just microevolution within a species." While this is true, the rapidity and obvious adaptiveness of the change effected by natural selection still seemed to give antievolutionists discomfort. Therefore, it is understandable that when Wells and his fans sniffed a scientific controversy over peppered moths (in truth it was a fairly marginal kind of controversy), they blew things way out of proportion.
- First, several of Wells's worst distortions must be dealt with directly.
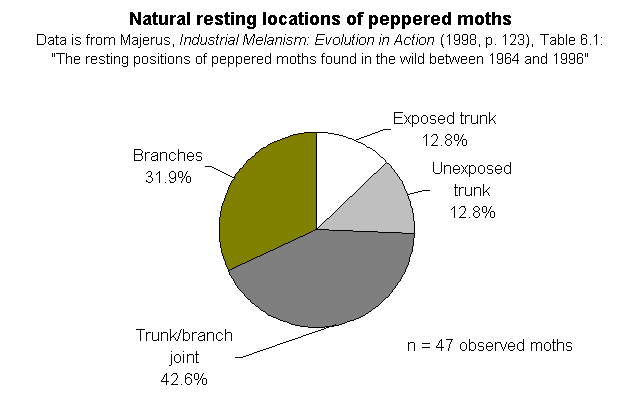
The natural resting locations of peppered moths -- Majerus' data. On page 148, Wells discusses the natural resting places of peppered moths, under the heading "Peppered moths don't rest on tree trunks." But they do, at least sometimes. Here are the relevant datasets, which Wells does not quote or cite for his readers:
For further discussion, see below and endnote 4.
Peppered moth photographs, staged and otherwise. Wells raises a fantastic stink about the fact that the photographs of peppered moths in textbooks, showing light-colored typicals next to dark-colored melanics on differing backgrounds, are staged. But the point of such photos is not to prove the truth of the 'classic' story, it is to illustrate the relative crypsis of moth morphs on different backgrounds. Those who feel that their innocent faith in insect photography has been betrayed should consider the fact that most photos of insects in textbooks are probably staged; insects are, after all, small and difficult to photograph. The facts that peppered moths are sparsely distributed and, well, camouflaged also make them difficult to photograph.
But as it turns out, the differences between staged and unstaged photos are minimal. Readers who wish to see unstaged photos of peppered moths are advised to look up Majerus' Melanism: Evolution in Action. Majerus says that all of the peppered moth photos taken by him in the book are unstaged. Readers should consult the figures which are listed below. It may be possible to get permission to include the photos, but until then descriptions shall have to suffice.
(For those with foggy memories of their texbooks, English peppered moths come in three general phenotypic categories: typica, the pale, original 'peppered' form of the moth; carbonaria, the almost black melanic form; and insularia, which includes a range of intermediate-colored moths.)
Figure 6.1 (a), p. 118. Black-and-white photo, edges blurred. A rather dark (almost black) insularia moth, resting apparently on a tree trunk (bark fills the background). The moth is slightly darker than the background.
Figure 6.1 (b), p. 118. Black-and-white photo, middle of moth slightly blurred. A light form of insularia (still more heavily peppered than a typica), resting on a thick tree branch (branch width is about 3/4 that of the moth).
Figure 6.3, p. 122. Black-and-white photo, middle of moth slightly blurred. A typica hanging underneath a hazel twig.
Plate 3, between pp. 146-147, has colored photos. Six photos are shown (the first five are Majerus'), and the captions are quoted, with my comments in brackets.
(a) "Typica and carbonaria forms of the peppered moth on an [sic] horizontal birch branch." [This situation, with two moths close enough together to photograph at once, is very rare, basically only occurring if two moths are meeting to mate.]
(b) "A pair of peppered moths on a twig at dawn. The carbonaria male is much less conspicuous than the typica female." [The carbonaria moth is quite blurry.]
(c) "A carbonaria peppered moth in shadow under a horizontal branch, showing how this positioning may reduce the likelihood of detection." [The moth is being viewed head-on and is indeed difficult to see.]
(d) "Typical form of the peppered moth at rest during the day in hazel foliage." [Head-on view, the moth is hanging underneath a thick twig.]
(e) "An intermediate, insularia form, of the peppered moth." [A 'classic' view, the moth is well-matched to its background, which is apparently tree bark.]
(f) "The non-melanic form of the peppered moth from North America, Biston betularia cognataria (courtesy of Professor Bruce Grant)." [A 'classic' view, the moth is well-matched to its background, which is a lichen-covered surface.]
It should be noted that Majerus is concerned to show his readers aspects of the peppered moth story that they do not get in textbooks; thus the focus on insularia forms and on moths in branches (Majerus is a proponent of the view that peppered moths most commonly -- but not entirely or even almost entirely -- rest on the underside of branches and thick twigs in the forest canopy). Even so, there are several photos that show peppered moths, on tree trunks, on more-or-less matching backgrounds. And guess what? These photos look no different than 'staged' photos of moths on tree-trunks. The most 'staged' aspect about a 'staged' photo is that two differing moth forms are shown side-by-side, but Majerus' first two photos from Plate 3 indicate that even this is not impossible. So the entire photo issue is a mountain made of a molehill.
It should also be noted that several (four) of these unstaged photos have some (minor but noticeable) degree of blurring (e.g., part of the moth will be out of focus). Insects in the wild do annoying things like move and fly away, and are often encountered in poor-light conditions, resulting in less-than-perfect photos. As scientific documentation of observations this is unimportant, but flawed photographs are exactly the kind of thing that are avoided in textbooks, and this is precisely why staging insect photos is a common practice for textbooks (as well as things like nature shows).
Summary of Wells's treatment of moth resting places. To review, Wells's primary objection to the peppered moth story was this:
Most introductory textbooks now illustrate this classical story of natural selection with photographs of the two varieties of peppered moth resting on light- and dark-colored tree trunks. (Figure 7-1) What the textbooks don't explain, however, is that biologists have known since the 1980's that the classical story has some serious flaws. The most serious is that peppered moths in the wild don't even rest on tree trunks. The textbook photographs, it turns out, have been staged. (Icons, p. 138)
[Figure 7-1 is on Icons, p. 139; these are drawings by Icons illustrator Jody F. Sjogren; the source photo, if there is one, is not cited. Confusingly, the caption for the figure is not on page 139 but overleaf on page 140. These are not encouraging signs in a book purporting to critique textbooks.]
The discussion thus far has shown that Wells's "most serious objection" to the peppered moth story is completely baseless: first, peppered moths do in fact rest on tree trunks (a significant portion of the time although not the majority of the time, according to Majerus' data). Second, textbook photos are used to show relative crypsis of moth morphs, not to prove that peppered moths always rest in one section of the trees. And third, Majerus himself has taken unstaged photos of peppered moths on matching tree trunk backgrounds, and these are not significantly different than staged photos; this eviscerates whatever vestige of a point Wells thinks that he has.
What are the implications if moths rest most often underneath branches? Leaving aside Wells's frantic attempt to create a problem where none exists, the relevance of moth resting locations for the 'classic story' (natural selection by bird predation) deserves some consideration. Majerus' considered opinion is that peppered moths rest more commonly underneath branches than was previously appreciated, and that if this is true then some quantitative estimates of selection coefficients may need to be adjusted. However, he is quite clear that the basic qualitative conclusions of Kettlewell (that differential bird predation of moth morphs on changing backgrounds is the selective force) do not need to be changed. As Majerus notes, crypsis is still important for moths in tree branches. He even comments directly on this with two of his photos (Plate 3, photos (b) and (c)). And of course, birds are known to (a) fly and (b) feed in forest canopies, so it is very difficult to see why resting on trunks vs. branches would change bird predation in any radical way.
The scientific literature. Having dealt with Wells's "most serious objection," let us turn to Wells's use of the scientific literature. The primary problem is that Wells gives inordinate weight to a few scattered review papers, by biologists who are not major peppered moth researchers [4], that question the standard view (that bird predation on different colored moths on differently polluted backgrounds caused the darkening of moth populations as pollution increased, and that as pollution decreased this process worked in the opposite direction). Their criticisms have been answered by peppered moth researchers (Grant, 1999; Cook, 2000; Grant and Clarke, 2000; Majerus, 2000). And, as pointed out in the introduction, since Wells bases his argument on the idea that the experts are disowning the 'icons' in their respective fields, Wells is falsified if those experts contradict him.
Bruce Grant's review of Wells. American peppered moth researcher Bruce Grant has written many papers on Biston, and has documented the parallel rise and fall in melanic forms of the North American subspecies of the peppered moth. See Grant's webpage [http://faculty.wm.edu/bsgran/] for listed articles. Dr. Grant has kindly given permission to have his comments on the peppered moth chapter of Icons quoted in this article.
To put them in context, the material quoted below is a copy of the correspondence between Grant and a professional colleague who had requested Grant's views on Wells' chapter, originally written February 7, 2001.
Subject: Wells's Chapter on Peppered Moths
Wells's Chapter 7 is pretty similar to his earlier ms. "Second thoughts about peppered moths" that he posted on the web, and published in abridged form in The Scientist. I sent you my comments about that version about two weeks ago. My general reaction to this latest version is about the same. He distorts the picture, but unfortunately he is probably pretty convincing to people who really don't know the primary literature in this field. He uses two tactics. One is the selective omission of relevant work. The other is to scramble together separate points so doubts about one carry over to the other. Basically, he is dishonest.
He immediately launches the claim "that peppered moths in the wild don't even rest on tree trunks" (p. 138). This is just plain wrong! Of course they rest on tree trunks, but it's not their exclusive resting site. He quotes Cyril Clarke's lack of success in finding the moths in natural settings, but he omits mentioning Majerus' data which reports just where on trees (exposed trunks, unexposed trunks, trunk/branch joints, branches) Majerus has found moths over his 34 years of looking for them. Of the 47 moths he located away from moth traps, 12 were on trunks (that's >25%). Of the 203 he found in the vicinities of traps, 70 were on trunks (that's 34%). Based on his observations, Majerus argued that the most common resting site appears to be at the trunk/branch juncture. What is clear from his data is that they sit all over the trees, INCLUDING the trunks. So what? Kettlewell's complementary experiments in polluted and unpolluted woods compared the relative success of different colored moths on the same parts of trees in different areas, not different parts of trees in the same area. It is true that the photos showing the moths on trunks are posed (just like practically all wildlife pictures of insects are) but they are not fakes. No one who reads Kettlewell's paper in which the original photos appeared would get the impression from the text that these were anything but posed pictures. He was attempting to compare the differences in conspicuousness of the pale and dark moths on different backgrounds. Nobody thought he encountered those moths like that in the wild. At their normal densities, you'd be hard pressed ever to find two together unless they were copulating. I have always made a point of stating in photo captions that the moths are posed, and I think textbook writers have been careless about this. But they are not frauds.
On the subject of lichens, no one has questioned their importance more than I have. But what does Wells do with this? He quotes me, but he doesn't include what else I said has happened on the Wirral (p. 147) with respect to the tremendous expansion of birch stands since the enactment of the smokeless zones. Kettlewell, too, argued that peppered moths are well concealed on birch bark (even without lichens). Wells continues (p. 148) to quote my reservations about lichens in Michigan, but, again, he omits any reference to the data I presented in that paper showing the decline, not only in SO2, but in atmospheric particles (soot) which has been established as a factor altering reflectance from the surface of tree bark. So, while I have questioned the importance of lichens, I have not taken this as evidence that crypsis is unimportant. Wells omits this entirely.
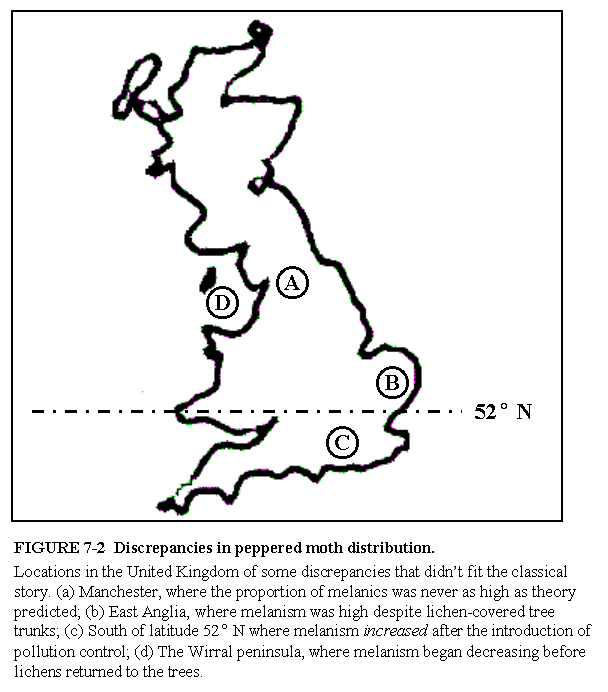
Wells continues to bring up the same old arguments about mysterious other factors (yet to be identified) that account for the persistence of typicals in polluted regions, and the presence of melanics in unpolluted locales. He cites papers written back in the 70s about these puzzles. He omits discussing in any sophisticated way the role of migration other than to say "Theoretical models could account for the discrepancies only by invoking migration...." (p.146), as if in desperation we are forced to grasp at straws. Of course migration is important. Majerus actually reviews this point fairly well by comparing the smoothness of clines in melanism between species that are highly mobile (as is Biston), and species that are relatively sedentary. Instead of showing his meaningless map of the UK (Fig. 7-2) to illustrate what he regards as anomalies in the distribution of melanism and lichens, why doesn't he show the before and after comparison from the national surveys by Kettlewell in 1956, and the survey by Grant et al. in 1996. (If you'd like, I can send you a jpg file of the maps I mean.)
Wells also inappropriately uses thermal melanism in ladybirds to suggest, that while no one has shown this in peppered moths (p. 152), industrial melanism can have other causes besides predation. It's not just that there is no evidence for thermal melanism in peppered moths, there is evidence AGAINST thermal melanism based on the geographic incidence of melanism in the UK, the USA, and Europe. There are no latitudinal clines, and no altitudinal clines as one might expect with thermal melanism. Wells knows this, if he actually read my papers. (He cites them, so I should assume he read them.) He also raises the question of larval tolerance to pollutants. There is no evidence for this, either. I have a paper out on this point, but in fairness to Wells, it came out just this past year.
Wells clouds discussions with irrelevancies. For examples he brings up Heslop Harrison (p. 141 and again on p. 151) and the question of phenotypic induction. Wells makes it sound as if most biologists discount induction based on their belief in natural selection (as if it were a popular religious question). The evidence for the Mendelian inheritance of melanism in peppered moths has nothing to do with evolutionary theory; it is based on old fashioned crosses involving over 12 thousand progeny from 83 broods. The Mendelian basis for this character in this species is as well established as is any character in any species. Wells doesn't mention this, yet he cites my review paper where I do bring this up in my criticism of Sargent et al. Induction has nothing to do with industrial melanism, and Wells knows it. Again, selective omissions on the part of Wells.
On page 151 Wells claims Kettlewell's evidence has been impeached. This is nonsense. It has not. But I have argued, that even if it were entirely thrown out, the evidence for natural selection comes from the changes in the percentages of pale and melanic moths. It is this record of change in allele frequency over time that is unimpeachable. It is a massive record by any standard. (I can send a jpg file with graphs, if you'd like.) I have pointed out, and he quotes me, that no force known to science can account for these changes except for natural selection. Yet, he scrambles the ingredients here. He claims (top of p. 153) "...it is clear that the compelling evidence for natural selection that biologists once thought they had in peppered moths no longer exists." Of course the evidence for natural selection exists! That evidence is overwhelming. Wells, by attempting to discredit Ketttlewell's experiments about predation (and clearly there are things wrong with Kettlewell's experiments) doesn't stop at saying we can't be altogether sure about bird predation because of problems with Kettlewell's experiments. No. He says, instead, that the evidence for natural selection no longer exists. This is just plain wrong. He cannot support this sweeping statement, but he spins it into his conclusion by building a case against Kettlewell. This is what I mean about his tactic of scrambling arguments. He wields non sequiturs relentlessly.
I hope this is helpful to you in your review.
Bruce Grant, Professor of Biology, College of William & Mary. February 2001
Michael Majerus' review of Wells. Majerus' views on the peppered moth "debate" were made clear in 1999, during an online fracas on the Calvin evolution listserv. Majerus, who as we have noted wrote the (most recent) book on peppered moths and industrial melanism (Majerus, 1998), was contacted by one of the participants, Don Frack, about the claims of creationists about the peppered moth now being a "peppered myth." The creationists' claims were theoretically based on Majerus' book, Coyne's (1998) review of it, and an article discussing these works and interviewing Majerus and Coyne in the British newspaper The Sunday Telegraph (Matthews, 1999). Majerus' email message was posted to the listserv. Majerus concluded:
Bernard [Kettlewell] was a first rate entomologist and scientist. His experiments were meticulous and generally well designed. In my opinion, many of his experiments were among the best that have been conducted on melanism and bird predation. The 'design flaws' in some of the experiments, if you want to call them that were primarily a result of practical expediency because Kettlewell wanted to be able to see birds taking moths, and to film them. The only real flaw may have been his resting site selection experiments, where he MIGHT (we do not actually know) have used moths from different populations (see pages 142-143).
[...]
The suggestion that Kettlewell ever 'faked' a result is offensive to his memory. He was an honourable, good scientist who reported his findings with honesty and integrity.
[...]
To end, may I put on record to you, that my view is that the rise and fall of the carbonaria form of the peppered moth has resulted from changes in the environments in which this moth lives. These changes have come about as a result on changes in pollution levels which have altered the relative crysis of the forms of this moth. The main, if not the only selective factor that has lead to changes in the frequencies of the forms over time is differential bird predation. The case of melanism in the peppered moth IS ONE OF THE BEST EXAMPLES OF EVOLUTION IN ACTION BY DARWIN'S PROCESS OF NATURAL SELECTION that we have. In general it is based on good science and it is sound.
(Majerus email to Don Frack, posted March 30, 1999. Capitalization original. Available at: http://www.calvin.edu/archive/evolution/199903/0312.html)
Wells was evidently contacted about Majerus' comments. Since Majerus had been Wells's primary source for the claim that the peppered moth was now a "peppered myth," perhaps Wells felt that he had to respond. Wells's response was to move Majerus from the "respected authority" category to the "fraud" category. Frankly, Wells freaks out:
BUT EVERYONE, INCLUDING MAJERUS, HAS KNOWN SINCE THE 1980'S THAT PEPPERED MOTHS DO NOT REST ON TREE TRUNKS IN THE WILD. This means that every time those staged photographs have been re-published since the 1980's constitutes a case of deliberate scientific fraud. Michael Majerus is being dishonest, and textbook-writers are lying to biology students. The behavior of these people is downright scandalous.
I know what I'm talking about. I spent much of last summer reading the primary literature (email me if you want the references). Frankly, I was shocked by what I found -- not only that the evidence for the moths' true resting-places has been known since the 1980's, but also that people like Majerus and Miller continue to deceive the public.
Fraud is fraud. It's time to tell it like it is.
(Wells's message posted to Calvin listserv, March 31, 1999. Capitalization original. Available at: http://www.asa3.org/archive/evolution/199903/0348.html)
In due course, Wells's message made it back to Majerus, who responded with a full-scale dissection of all of Wells's key points:
[...]
Evidence of selective predation in the peppered moth is not lacking. It is just not provided in the quick text book descriptions of the peppered moth. How can it be. I have read some 500 papers on melanism in the Lepidoptera. In total, these papers probably amount to about 8000 pages, and the story is condensed into a few paragraphs in most textbooks for schools. Even in my own book, I could only give a review of the case covering about 60 pages including illustrations.
The older hypothesis that melanism was induced by pollutants was discredited because Heslop [Harrison's] experiments lacked appropriate controls, and his results could not be replicated, despite several attempts. Furthermore, the levels of mutagenesis that he recorded are several times higher than those produced by doses of radiation that induce complete sterility in fruit flies (see E.B. Ford (1964) Ecological Genetics for full critical review).
Finally, I agree with Dr Wells that photographs of two peppered moths staged on backgrounds for effect should say they have been done purely for illustrative purposes. I have many times, in undergraduate lectures, pointed out that photographs of the type that appear in so many text books are faked. However, I would point out that none of the photographs of live peppered moths taken by myself, which appear in the book were staged. All show peppered moths where they were found in the wild.
End-note: It is difficult to have an informed discussion of a complicated ecological system with those who have little or no experience of the system. My advice to anyone who wishes to obtain a fully objective view of this case is to a) read the primary papers that I based my review upon, and any other relevant papers, and b) gain some experience of this moth and its habits in the wild. Of all the people I know, including both amateur and professional entomologists who have experience of this moth, I know of none who doubts that differential bird predation is of primary importance in the spread and decline of melanism in the peppered moth.
I hope that this is some use to you, Donald, and that it encourages more people to look at the case of the peppered moth with an open mind. If it can help interest a few more people in moths and butterflies, that is all to the good.
Best wishes, and Happy Easter.
Mike Majerus
(Majerus email, posted to Calvin listserv by Don Frack, April 5, 1999, bold added. Available at: http://www.asa3.org/archive/evolution/199904/0103.html)Frack says of this,
Note the complete irony of the capitalized sentence. Majerus is the foremost proponent (in the literature I've seen) of the idea that the moths most commonly rest higher in the trees. His data are the only ones I have seen cited as evidence of [what] happens "in the wild." Majerus is attacked as "dishonest" and "text-book writers are lying to biology students", their behavior is "scandalous." [...] If Wells is right, he hasn't demonstrated it here. He attacks both Michael Majerus and Bruce Grant. If Grant's frequent co-authors, such as Cyril Clarke, are added to the ridicule list (and I don't know why they wouldn't be), then Wells is well on his way to rejecting all the well-known researchers on this subject. An awesome, and, at face value, an incredibly arrogant, claim.
There is much more where this came from, and unfortunately there is not yet a comprehensive web source covering Wells's abuse of peppered moths, so you have to do some digging. Some good places to start are these links:
Bruce Grant's review article: http://www.wm.edu/biology/melanism.pdf
Don Lindsay's archive (links to various letters to newspapers from Grant and others, protesting Wells's characterizations of their work)
Wells scored a minor coup when a toned-down version of his essay "Second Thoughts about Peppered Moths" was published in The Scientist (13(11), p. 13, May 24, 1999). A longer, unedited version is here.
The 1999 Wells/Frack posts on the Calvin listserv go roughly in this order (these really should be reformatted and archived somewhere):
Frack, "Peppered Moths - in black and white (part 1 of 2)":
http://www.asa3.org/archive/evolution/199903/0314.htmlFrack, "Peppered Moths - in black and white (part 2 of 2)":
http://www.asa3.org/archive/evolution/199903/0312.htmlWells, quoted in "Peppered moths again":
http://www.asa3.org/archive/evolution/199903/0348.htmlFrack, "RE: Peppered Moths again": http://www.asa3.org/archive/evolution/199903/0378.html
Frack, Peppered Moths - round 2 (part 1 of 2)
http://www.asa3.org/archive/evolution/199904/0100.htmlFrack, Peppered Moths - round 2 (part 2 of 2)
http://www.asa3.org/archive/evolution/199904/0103.htmlFrack, "Peppered moths, round 3":
http://www.asa3.org/archive/evolution/199904/0200.htmlFrack, "Peppered moths and Creationists":
http://www.asa3.org/archive/evolution/199904/0201.htmlWells, "My last word":
http://www.asa3.org/archive/evolution/199904/0204.htmlFrack, "RE: My last word":
http://www.asa3.org/archive/evolution/199904/0207.htmlOf moths and maps. So, the experts disagree with Wells and furthermore identify just the kinds of deceptive tactics that I have been talking about. A further instance is Wells's Figure 7-2 (p. 145), a map of England with four locations marked with letters representing "Discrepancies in peppered moth distribution." This map deserves enshrinement as exhibit A in geographer Mark Monmonier's (1996) book How to Lie with Maps. This is basically what Wells's map looks like:
(After Wells, Icons, p. 145, Figure 7-2. The exact caption is quoted below the figure. My rendition of the border of Great Britain is very crude, being based on the first graphic I could find on the internet, but apart from that Wells's figure is accurately represented. A figure similar to that in Icons can be found in Wells's unedited moth essay, here.)
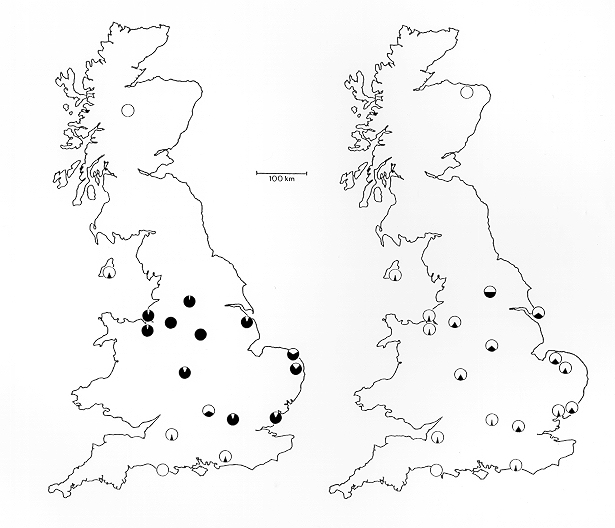
What Wells did here was dig through the literature and find a few instances where he could find some weak excuse for an observation 'contradicting' what was expected. However, if one inspects real maps by real moth researchers, one finds that the geographic pattern is actually a good match with expectations. Here are the maps that Bruce Grant mentioned in the above-quoted review:
Grant's comments on these maps: "The maps show a before-after comparison of the geographic distribution of melanic phenotypes in peppered moth populations in Britain based on Kettlewell's 1956 survey (left map) and that conducted 40-years later (1996) by my colleagues and me (right map). The black segments of the pie charts indicate the percentage of melanics at the various locations. Clearly melanism has declined everywhere it was once common." (Grant, personal communication, February 11, 2002)
The source publication for these maps: Grant, B. S., Cook, A. D. , Clarke, C. A., and Owen, D. F. 1998. Geographic and temporal variation in the incidence of melanism in peppered moth populations in America and Britain. Journal of Heredity 89:465-471.
Wells's map (Figure 7-2 from Icons) is more fraudulent than all of the textbook moth photos put together.
You bet, if you're going to make something your lifes work, you may as well be good at it. :-}
I'm not against evolution. I'm against the darwinite religion. You're telling me running software on a computer requires no intelligent design! You probably believe Bill Gates was just lucky. Time, Chance and Miracles. You will not brainwash me into your sect.
Modern "evolving code" project: Avida
Put this down under *Just Damn*. Scooped again!
I was wondering ten years ago why people didn't do this.
As far as its relevance to biological evolution, your disputant will point out to the (however many) non-functional copies and say "See? That's what I said!" because what they had in mind was random mutations within a single copy.
You wouldn't mind helping me invest in lottery tickets so I'd have enough time to read this, the earlier articles on sulphide hotspots, etc. etc., would you? :-)
Peppered moths and finches....the best evidence of a grandiose myth
Then why do you keep objecting to it?
I'm against the darwinite religion.
If you ever find something resembling a "darwinite religion", do let me know. In the meantime I'll keep discussing evolutionary biology.
You're telling me running software on a computer requires no intelligent design!
Reading comprehension isn't your strong suit, is it? No, that's not what I'm saying. I was actually quite clear -- try reading my post again, this time for content.
You probably believe Bill Gates was just lucky. Time, Chance and Miracles.
Ramble much?
You will not brainwash me into your sect.
Ooookay...
The astute reader is invited to examine post #158, and decide for himself whether "peppered moths and finches" are the "best evidence" for evolution, or whether there's a ton of really solid evidence for it which "metacognative" has seen but keeps pretending he hasn't...
I'd like to think that not all anti-evolutionists are such classic examples of "there are none so blind as he who will not see", but that's not been my experience with them.
Placemarker.
I betcha the sentiment he had in mind was more or less what you expressed later in these lines:
I've *DONE* genetic algorithms. I *know* that I constructed the "experiment"
e.g. someone had to write the original source, compile it, and (presumably) hit "Run" or whatever to start things off.
BTW, as a designer of the code, what did you have for dinner?
Disclaimer: Opinions posted on Free Republic are those of the individual posters and do not necessarily represent the opinion of Free Republic or its management. All materials posted herein are protected by copyright law and the exemption for fair use of copyrighted works.