Skip to comments.
Life's Irreducible Structure (DEBATE THREAD)
CMI ^
| Alex Williams
Posted on 01/12/2009 7:23:26 AM PST by GodGunsGuts
DEBATE THREAD
This INFORMAL debate will focus on Part 1 and 2 of Alex Williams' paper "Life's irreducible structure." Williams' paper will serve as the affirmative, namely:
(A) All aspects of life (not just bacterial flagellums and blood clotting cascades) lie beyond the reach of naturalistic explanations, and (B) only intelligent design meets the criterion of an acceptable historical inference according to the Law of Cause and Effect.
Part 1 of Alex Williams' paper follows. A link to Part 2 can be found in reply #1. It is strongly suggested that both papers be read before participating in the discussion/debate.
One final note: please refrain from making rude comments, please try to ignore those who do, and please try to stay on topic. All the best--GGG
-----------------------------------------------------------------------
Life’s irreducible structure—Part 1: autopoiesis
by Alex Williams
The commonly cited case for intelligent design appeals to: (a) the irreducible complexity of (b) some aspects of life. But complex arguments invite complex refutations (valid or otherwise), and the claim that only some aspects of life are irreducibly complex implies that others are not, and so the average person remains unconvinced. Here I use another principle—autopoiesis (self-making)—to show that all aspects of life lie beyond the reach of naturalistic explanations. Autopoiesis provides a compelling case for intelligent design in three stages: (i) autopoiesis is universal in all living things, which makes it a pre-requisite for life, not an end product of natural selection; (ii) the inversely-causal, information-driven, structured hierarchy of autopoiesis is not reducible to the laws of physics and chemistry; and (iii) there is an unbridgeable abyss between the dirty, mass-action chemistry of the natural environmental and the perfectly-pure, single-molecule precision of biochemistry. Naturalistic objections to these propositions are considered in Part II of this article.
Snowflake photos by Kenneth G. Libbrecht.
Figure 1. Reducible structure. Snowflakes (left) occur in hexagonal shapes because water crystallizes into ice in a hexagonal pattern (right). Snowflake structure can therefore be reduced to (explained in terms of) ice crystal structure. Crystal formation is spontaneous in a cooling environment. The energetic vapour molecules are locked into solid bonds with the release of heat to the environment, thus increasing overall entropy in accord with the second law of thermodynamics.
The commonly cited case for intelligent design (ID) goes as follows: ‘some biological systems are so complex that they can only function when all of their components are present, so that the system could not have evolved from a simpler assemblage that did not contain the full machinery.’1 This definition is what biochemist Michael Behe called irreducible complexity in his popular book Darwin’s Black Box2 where he pointed to examples such as the blood-clotting cascade and the proton-driven molecular motor in the bacterial flagellum. However, because Behe appealed to complexity, many equally complex rebuttals have been put forward,3 and because he claimed that only some of the aspects of life were irreducibly complex, he thereby implied that the majority of living structure was open to naturalistic explanation. As a result of these two factors, the concept of intelligent design remains controversial and unproven in popular understanding.
In this article, I shall argue that all aspects of life point to intelligent design, based on what European polymath Professor Michael Polanyi FRS, in his 1968 article in Science called ‘Life’s Irreducible Structure.’4 Polanyi argued that living organisms have a machine-like structure that cannot be explained by (or reduced to) the physics and chemistry of the molecules of which they consist. This concept is simpler, and broader in its application, than Behe’s concept of irreducible complexity, and it applies to all of life, not just to some of it.
The nature and origin of biological design
Biologists universally admire the wonder of the beautiful ‘designs’ evident in living organisms, and they often recoil in revulsion at the horrible ‘designs’ exhibited by parasites and predators in ensuring the survival of themselves and their species. But to a Darwinist, these are only ‘apparent designs’—the end result of millions of years of tinkering by mutation and fine tuning by natural selection. They do not point to a cosmic Designer, only to a long and ‘blind’ process of survival of the fittest.5 For a Darwinist, the same must also apply to the origin of life—it must be an emergent property of matter. An emergent property of a system is some special arrangement that is not usually observed, but may arise through natural causes under the right environmental conditions. For example, the vortex of a tornado is an emergent property of atmospheric movements and temperature gradients. Accordingly, evolutionists seek endlessly for those special environmental conditions that may have launched the first round of carbon-based macromolecules6 on their long journey towards life. Should they ever find those unique environmental conditions, they would then be able to explain life in terms of physics and chemistry. That is, life could then be reduced to the known laws of physics, chemistry and environmental conditions.
However, Polanyi argued that the form and function of the various parts of living organisms cannot be reduced to (or explained in terms of) the laws of physics and chemistry, and so life exhibits irreducible structure. He did not speculate on the origin of life, arguing only that scientists should be willing to recognize the impossible when they see it:
‘The recognition of certain basic impossibilities has laid the foundations of some major principles of physics and chemistry; similarly, recognition of the impossibility of understanding living things in terms of physics and chemistry, far from setting limits to our understanding of life, will guide it in the right direction.’7
Reducible and irreducible structures
To understand Polanyi’s concept of irreducible structure, we must first look at reducible structure. The snowflakes in figure 1 illustrate reducible structure.
Meteorologists have recognized about eighty different basic snowflake shapes, and subtle variations on these themes add to the mix to produce a virtually infinite variety of actual shapes. Yet they all arise from just one kind of molecule—water. How is this possible?
Figure 2. Irreducible structure. The silver coins (left) have properties of flatness, roundness and impressions on faces and rims, that cannot be explained in terms of the crystalline state of silver (close packed cubes) or its natural occurrence as native silver (right).
When water freezes, its crystals take the form of a hexagonal prism. Crystals then grow by joining prism to prism. The elaborate branching patterns of snowflakes arise from the statistical fact that a molecule of water vapour in the air is most likely to join up to its nearest surface. Any protruding bump will thus tend to grow more quickly than the surrounding crystal area because it will be the nearest surface to the most vapour molecules.8 There are six ‘bumps’ (corners) on a hexagonal prism, so growth will occur most rapidly from these, producing the observed six-armed pattern.
Snowflakes have a reducible structure because you can produce them with a little bit of vapour or with a lot. They can be large or small. Any one water molecule is as good as any other water molecule in forming them. Nothing goes wrong if you add or subtract one or more water molecules from them. You can build them up one step at a time, using any and every available water molecule. The patterns can thus all be explained by (reduced to) the physics and chemistry of water and the atmospheric conditions.
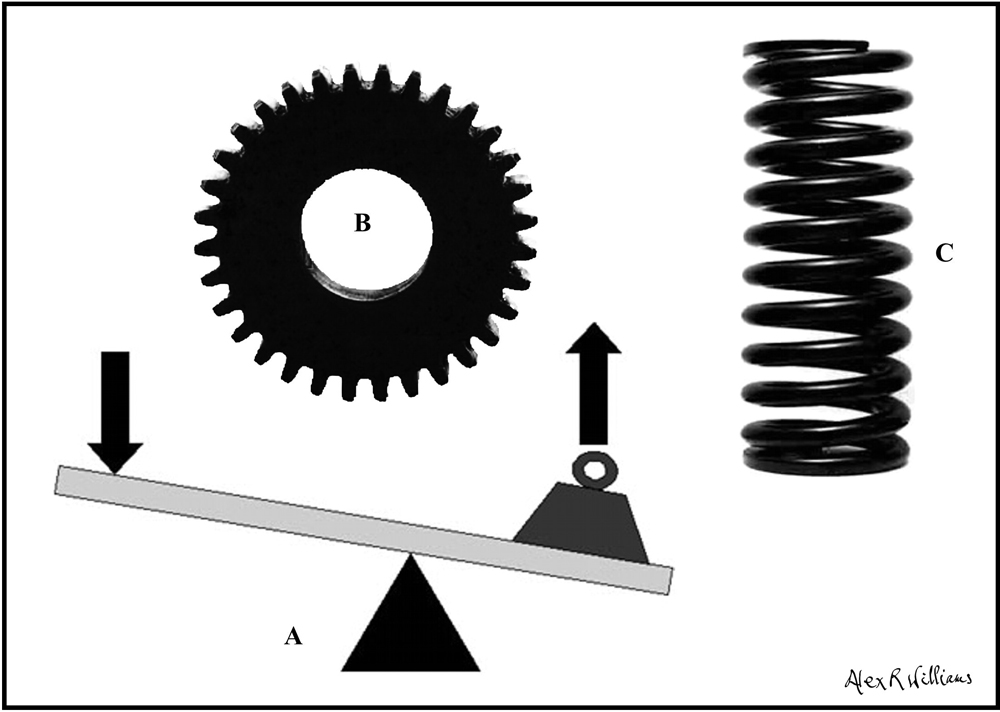
Figure 3. Common irreducibly structured machine components: lever (A), cogwheel (B) and coiled spring (C). All are made of metal, but their detailed structure and function cannot be reduced to (explained by) the properties of the metal they are made of.
To now understand irreducible structure, consider a silver coin.
Silver is found naturally in copper, lead, zinc, nickel and gold ores—and rarely, in an almost pure form called ‘native silver’. Figure 2 shows the back and front of two vintage silver coins, together with a nugget of the rare native form of silver. The crystal structure of solid silver consists of closely packed cubes. The main body of the native silver nugget has the familiar lustre of the pure metal, and it has taken on a shape that reflects the available space when it was precipitated from groundwater solution. The black encrustations are very fine crystals of silver that continued to grow when the rate of deposition diminished after the main load of silver had been deposited out of solution.
Unlike the case of the beautifully structured snowflakes, there is no natural process here that could turn the closely packed cubes of solid silver into round, flat discs with images of men, animals and writing on them. Adding more or less silver cannot produce the roundness, flatness and image-bearing properties of the coins, and looking for special environmental conditions would be futile because we recognize that the patterns are man-made. The coin structure is therefore irreducible to the physics and chemistry of silver, and was clearly imposed upon the silver by some intelligent external agent (in this case, humans).
Whatever the explanation, however, the irreducibility of the coin structure to the properties of its component silver constitutes what I shall call a ‘Polanyi impossibility’. That is, Polanyi identified this kind of irreducibility as a naturalistic impossibility, and argued that it should be recognized as such by the scientific community, so I am simply attaching his name to the principle.
Polanyi pointed to the machine-like structures that exist in living organisms. Figure 3 gives three examples of common machine components: a lever, a cogwheel and a coiled spring. Just as the structure and function of these common machine components cannot be explained in terms of the metal they are made of, so the structure and function of the parallel components in life cannot be reduced to the properties of the carbon, hydrogen, oxygen, nitrogen, phosphorus, sulphur and trace elements that they are made of. There are endless examples of such irreducible structures in living systems, but they all work under a unifying principle called ‘autopoiesis’.
Autopoiesis defined
Autopoiesis literally means ‘self-making’ (from the Greek auto for self, and the verb poiéō meaning ‘I make’ or ‘I do’) and it refers to the unique ability of a living organism to continually repair and maintain itself—ultimately to the point of reproducing itself—using energy and raw materials from its environment. In contrast, an allopoietic system (from the Greek allo for other) such as a car factory, uses energy and raw materials to produce an organized structure (a car) which is something other than itself (a factory).9
Autopoiesis is a unique and amazing property of life—there is nothing else like it in the known universe. It is made up of a hierarchy of irreducibly structured levels. These include: (i) components with perfectly pure composition, (ii) components with highly specific structure, (iii) components that are functionally integrated, (iv) comprehensively regulated information-driven processes, and (v) inversely-causal meta-informational strategies for individual and species survival (these terms will be explained shortly). Each level is built upon, but cannot be explained in terms of, the level below it. And between the base level (perfectly pure composition) and the natural environment, there is an unbridgeable abyss. The enormously complex details are still beyond our current knowledge and understanding, but I will illustrate the main points using an analogy with a vacuum cleaner.
A vacuum cleaner analogy
My mother was excited when my father bought our first electric vacuum cleaner in 1953. It consisted of a motor and housing, exhaust fan, dust bag, and a flexible hose with various end pieces. Our current machine uses a cyclone filter and follows me around on two wheels rather than on sliders as did my mother’s original one. My next version might be the small robotic machine that runs around the room all by itself until its battery runs out. If I could afford it, perhaps I might buy the more expensive version that automatically senses battery run-down and returns to its induction housing for battery recharge.
Notice the hierarchy of control systems here. The original machine required an operator and some physical effort to pull the machine in the required direction. The transition to two wheels allows the machine to trail behind the operator with little effort, and the cyclone filter eliminates the messy dust bag. The next transition to on-board robotic control requires no effort at all by the operator, except to initiate the action to begin with and to take the machine back to the power source for recharge when it has run down. And the next transition to automatic sensing of power run-down and return-to-base control mechanism requires no effort at all by the operator once the initial program is set up to tell the machine when to do its work.
If we now continue this analogy to reach the living condition of autopoiesis, the next step would be to install an on-board power generation system that could use various organic, chemical or light sources from the environment as raw material. Next, install a sensory and information processing system that could determine the state of both the external and internal environments (the dirtiness of the floor and the condition of the vacuum cleaner) and make decisions about where to expend effort and how to avoid hazards, but within the operating range of the available resources. Then, finally, the pièce de résistance, to install a meta-information (information about information) facility with the ability to automatically maintain and repair the life system, including the almost miraculous ability to reproduce itself—autopoiesis.
Notice that each level of structure within the autopoietic hierarchy depends upon the level below it, but it cannot be explained in terms of that lower level. For example, the transition from out-sourced to on-board power generation depends upon their being an electric motor to run. An electric vacuum cleaner could sit in the cupboard forever without being able to rid itself of its dependence upon an outside source of power—it must be imposed from the level above, for it cannot come from the level below. Likewise, autopoiesis is useless if there is no vacuum cleaner to repair, maintain and reproduce. A vacuum cleaner without autopoietic capability could sit in the cupboard forever without ever attaining to the autopoietic stage—it must be imposed from the level above, as it cannot come from the level below.
The autopoietic hierarchy is therefore structured in such a way that any kind of naturalistic transition from one level to a higher level would constitute a Polanyi impossibility. That is, the structure at level i is dependent upon the structure at level i-1 but cannot be explained by the structure at that level. So the structure at level i must have been imposed from level i or above.
The naturalistic abyss
Most origin-of-life researchers agree (at least in the more revealing parts of their writings)10 that there is no naturalistic experimental evidence directly demonstrating a pathway from non-life to life. They continue their research, however, believing that it is just a matter of time before we discover that pathway. But by using the vacuum cleaner analogy, we can give a solid demonstration that the problem is a Polanyi impossibility right at the foundation—life is separated from non-life by an unbridgeable abyss.
Dirty, mass-action environmental chemistry
The ‘simple’ structure of the early vacuum cleaner is not simple at all. It is made of high-purity materials (aluminium, plastic, fabric, copper wire, steel plates etc) that are specifically structured for the job in hand and functionally integrated to achieve the designed task of sucking up dirt from the floor. Surprisingly, the dirt that it sucks up contains largely the same materials that the vacuum cleaner itself is made of—aluminium, iron and copper in the mineral grains of dirt, fabric fibres in the dust, and organic compounds in the varied debris of everyday home life. However, it is the difference in form and function of these otherwise similar materials that distinguishes the vacuum cleaner from the dirt on the floor. In the same way, it is the amazing form and function of life in a cell that separates it from the non-life in its environment.
Naturalistic chemistry is invariably ‘dirty chemistry’ while life uses only ‘perfectly-pure chemistry’. I have chosen the word ‘dirty chemistry’ not in order to denigrate origin-of-life research, but because it is the term used by Nobel Prize winner Professor Christian de Duve, a leading atheist researcher in this field.11 Raw materials in the environment, such as air, water and soil, are invariably mixtures of many different chemicals. In ‘dirty chemistry’ experiments, contaminants are always present and cause annoying side reactions that spoil the hoped-for outcomes. As a result, researchers often tend to fudge the outcome by using artificially purified reagents. But even when given pure reagents to start with, naturalistic experiments typically produce what a recent evolutionist reviewer variously called ‘muck’, ‘goo’ and ‘gunk’12—which is actually toxic sludge. Even our best industrial chemical processes can only produce reagent purities in the order of 99.99%. To produce 100% purity in the laboratory requires very highly specialized equipment that can sort out single molecules from one another.
Another crucial difference between environmental chemistry and life is that chemical reactions in a test tube follow the Law of Mass Action.13 Large numbers of molecules are involved, and the rate of a reaction, together with its final outcome, can be predicted by assuming that each molecule behaves independently and each of the reactants has the same probability of interacting. In contrast, cells metabolize their reactants with single-molecule precision, and they control the rate and outcome of reactions, using enzymes and nano-scale-structured pathways, so that the result of a biochemical reaction can be totally different to that predicted by the Law of Mass Action.
The autopoietic hierarchy
Perfectly-pure, single-molecule-specific bio-chemistry
The vacuum cleaner analogy breaks down before we get anywhere near life because the chemical composition of its components is nowhere near pure enough for life. The materials suitable for use in a vacuum cleaner can tolerate several percent of impurities and still produce adequate performance, but nothing less than 100% purity will work in the molecular machinery of the cell.
One of the most famous examples is homochirality. Many carbon-based molecules have a property called ‘chirality’—they can exist in two forms that are mirror images of each other (like our left and right hands) called ‘enantiomers’. Living organisms generally use only one of these enantiomers (e.g. left-handed amino acids and right-handed sugars). In contrast, naturalistic experiments that produce amino acids and sugars always produce an approximately 50:50 mixture (called a ‘racemic’ mixture) of the left-and right-handed forms. The horrors of the thalidomide drug disaster resulted from this problem of chirality. The homochiral form of one kind had therapeutic benefits for pregnant women, but the other form caused shocking fetal abnormalities.
The property of life that allows it to create such perfectly pure chemical components is its ability to manipulate single molecules one at a time. The assembly of proteins in ribosomes illustrates this single-molecule precision. The recipe for the protein structure is coded onto the DNA molecule. This is transcribed onto a messenger-RNA molecule which then takes it to a ribosome where a procession of transfer-RNA molecules each bring a single molecule of the next required amino acid for the ribosome to add on to the growing chain. The protein is built up one molecule at a time, and so the composition can be monitored and corrected if even a single error is made.
Specially structured molecules
Life contains such a vast new world of molecular amazement that no one has yet plumbed the depths of it. We cannot hope to cover even a fraction of its wonders in a short article, so I will choose just one example. Proteins consist of long chains of amino acids linked together. There are 20 amino acids coded for in DNA, and proteins commonly contain hundreds or even thousands of amino acids. Cyclin B is an averaged-size protein, with 433 amino acids. It belongs to the ‘hedgehog’ group of signalling pathways which are essential for development in all metazoans. Now there are 20433 (20 multiplied by itself 433 times) = 10563 (10 multiplied by itself 563 times) possible proteins that could be made from an arbitrary arrangement of 20 different kinds of amino acids in a chain of 433 units. The human body—the most complex known organism—contains somewhere between 105 (= 100,000) and 106 (=1,000,000) different proteins. So the probability (p) that an average-sized biologically useful protein could arise by a chance combination of 20 different amino acids is about p = 106 /10563 = 1/10557 . And this assumes that only L-amino acids are being used—i.e. perfect enantiomer purity.14
For comparison, the chance of winning the lottery is about 1/106 per trial, and the chance of finding a needle in a haystack is about 1/1011 per trial. Even the whole universe only contains about 1080 atoms, so there are not even enough atoms to ensure the chance assembly of even a single average-sized biologically useful molecule. Out of all possible proteins, those we see in life are very highly specialized—they can do things that are naturally not possible. For example, some enzymes can do in one second what natural processes would take a billion years to do.15 Just like the needle in the haystack. Out of all the infinite possible arrangements of iron alloy (steel) particles, only those with a long narrow shape, pointed at one end and with an eye-loop at the other end, will function as a needle. This structure does not arise from the properties of steel, but is imposed from outside.
Water, water, everywhere
There is an amazing paradox at the heart of biology. Water is essential to life,16 but also toxic—it splits up polymers by a process called hydrolysis, and that is why we use it to wash with. Hydrolysis is a constant hazard to origin-of-life experiments, but it is never a problem in cells, even though cells are mostly water (typically 60–90%). In fact, special enzymes called hydrolases are required in order to get hydrolysis to occur at all in a cell.17 Why the difference? Water in a test tube is free and active, but water in cells is highly structured, via a process called ‘hydrogen bonding’, and this water-structure is comprehensively integrated with both the structure and function of all the cell’s macromolecules:
‘The hydrogen-bonding properties of water are crucial to [its] versatility, as they allow water to execute an intricate three-dimensional “ballet”, exchanging partners while retaining complex order and enduring effects. Water can generate small active clusters and macroscopic assemblies, which can both transmit and receive information on different scales.’18
Water should actually be first on the list of molecules that need to be specially configured for life to function. Both the vast variety of specially structured macromolecules and their complementary hydrogen-bonded water structures are required at the same time. No origin-of-life experiment has ever addressed this problem.
Functionally integrated molecular machines
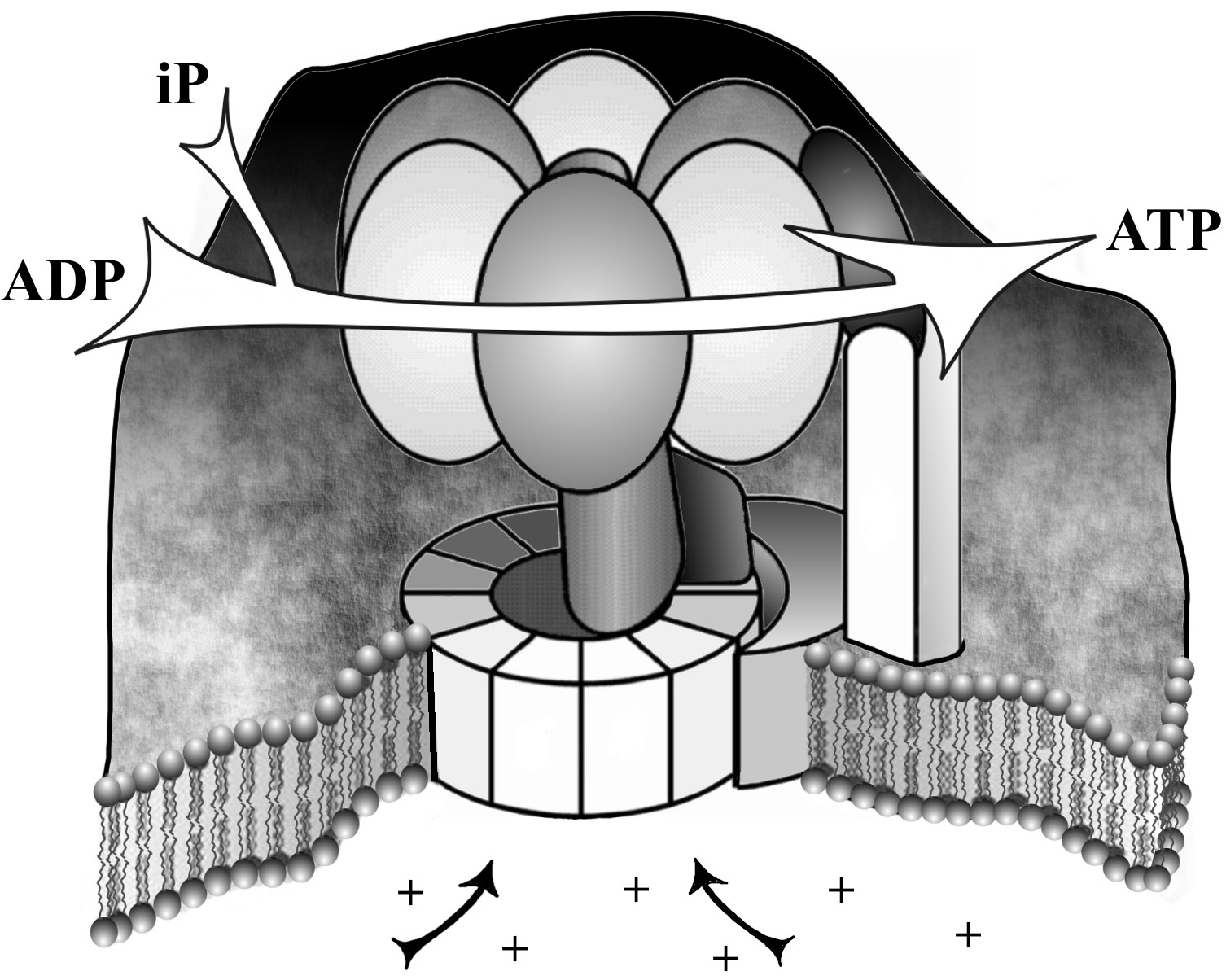
Figure 4. ATP synthase, a proton-powered molecular motor. Protons (+) from inside the cell (below) move through the stator mechanism embedded in the cell membrane and turn the rotor (top part) which adds inorganic phosphate (iP) to ADP to convert it to the high-energy state ATP.
It is not enough to have specifically structured, ultra-pure molecules, they must also be integrated together into useful machinery. A can of stewed fruit is fully of chemically pure and biologically useful molecules but it will never produce a living organism19 because the molecules have been disorganized in the cooking process. Cells contain an enormous array of useful molecular machinery. The average machine in a yeast cell contains 5 component proteins,20 and the most complex—the spliceosome, that orchestrates the reading of separated sections of genes—consists of about 300 proteins and several nucleic acids.21
One of the more spectacular machines is the tiny proton-powered motor that produces the universal energy molecule ATP (adenosine tri-phosphate) illustrated in Figure 4. When the motor spins one way, it takes energy from digested food and converts it into the high-energy ATP, and when the motor spins the other way, it breaks down the ATP in such a way that its energy is available for use by other metabolic processes.22
Comprehensively regulated, information-driven metabolic functions
It is still not enough to have spectacular molecular machinery—the various machines must be linked up into metabolic pathways and cycles that work towards an overall purpose. What purpose? This question is potentially far deeper than science can take us, but science certainly can ascertain that the immediate practical purpose of the amazing array of life structures is the survival of the individual and perpetuation of its species.23 Although we are still unravelling the way cells work, a good idea of the multiplicity of metabolic pathways and cycles can be found in the BioCyc collection. The majority of organisms so far examined, from microbes to humans, have between 1,000 and 10,000 different metabolic pathways.24 Nothing ever happens on its own in a cell—something else always causes it, links with it or benefits or is affected by it. And all of these links are multi-step processes.
All of these links are also ‘choreographed’ by information—a phenomenon that never occurs in the natural environment. At the bottom of the information hierarchy is the storage molecule—DNA. The double-helix of DNA is ‘just right’ for genetic information storage, and this ‘just right’ structure is beautifully matched by the elegance and efficiency of the code in which the cell’s information is written there.25 But it is not enough even to have an elegant ‘just right’ information storage system—it must also contain information. And not just biologically relevant information, but brilliantly inventive strategies and tactics to guide living things through the extraordinary challenges they face in their seemingly miraculous achievements of metabolism and reproduction. Yet even ingenious strategies and tactics are not enough. Choreography requires an intricate and harmonious regulation of every aspect of life to make sure that the right things happen at the right time, and in the right sequence, otherwise chaos and death soon follow.
Recent discoveries show that biochemical molecules are constantly moving, and much of their amazing achievements are the result of choreographing all this constant and complex movement to accomplish things that static molecules could never achieve. Yet there is no spacious ‘dance floor’ on which to choreograph the intense and lightning-fast (up to a million events per second for a single reaction26) activity of metabolism. A cell is more like a crowded dressing room than a dance floor, and in a show with a cast of millions!
Inversely causal meta-information
The Law of Cause and Effect is one of the most fundamental in all of science. Every scientific experiment is based upon the assumption that the end result of the experiment will be caused by something that happens during the experiment. If the experimenter is clever enough, then he/she might be able to identify that cause and describe how it produced that particular result or effect.
Causality always happens in a very specific order—the cause always comes before the effect.27 That is, event A must always precede event B if A is to be considered as a possible cause of B. If we happened to observe that A occurred after B, then this would rule out A as a possible cause of B.
In living systems however, we see the universal occurrence of inverse causality. That is, an event A is the cause of event B, but A exists or occurs after B. It is easier to understand the biological situation if we refer to examples from human affairs. In economics, for example, it occurs when behaviour now, such as an investment decision, is influenced by some future event, such as an anticipated profit or loss. In psychology, a condition that exists now, such as anxiety or paranoia, may be caused by some anticipated future event, such as harm to one’s person. In the field of occupational health and safety, workplace and environmental hazards can exert direct toxic effects upon workers (normal causality), but the anticipation or fear of potential future harm can also have an independently toxic effect (inverse causality).
Darwinian philosopher of science Michael Ruse recently noted that inverse causality is a universal feature of life,28 and his example was that stegosaur plates begin forming in the embryo but only have a function in the adult—supposedly for temperature control. However most biologists avoid admitting such things because it suggests that life might have purpose (a future goal), and this is strictly forbidden to materialists.
The most important example of inverse causality in living organisms is, of course, autopoiesis. We still do not fully understand it, but we do understand the most important aspects. Fundamentally, it is meta-information—it is information about information. It is the information that you need to have in order to keep the information you want to have to stay alive, and to ensure the survival of your descendants and the perpetuation of your species.
This last statement is the crux of this whole paper, so to illustrate its validity lets go back to the vacuum cleaner analogy. Let’s imagine that one lineage of vacuum cleaners managed to reach the robotic, energy-independent stage, but lacked autopoiesis, while a second makes it all the way to autopoiesis. What is the difference between these vacuum cleaners? Both will function very well for a time. But as the Second Law of Thermodynamics begins to take its toll, components will begin to wear out, vibrations will loosen connections, dust will gather and short circuit the electronics, blockages in the suction passage will reduce cleaning efficiency, wheel axles will go rusty and make movement difficult, and so on. The former will eventually die and leave no descendants. The latter will repair itself, keep its components running smoothly and reproduce itself to ensure the perpetuation of its species.
But what happens if the environment changes and endangers the often-delicate metabolic cycles that real organisms depend upon? Differential reproduction is the solution. Evolutionists from Darwin to Dawkins have taken this amazing ability for granted, but it cannot be overlooked. There are elaborate systems in place—for example, the diploid to haploid transition in meiosis, the often extraordinary embellishments and rituals of sexual encounters, the huge number of permutations and combinations provided for in recombination mechanisms—to provide offspring with variations from their parents that might prove of survival value. To complement these potentially dangerous deviations from the tried-and-true there are also firm conservation measures in place to protect the essential processes of life (e.g. the ability to read the DNA code and to translate it into metabolic action). None of this should ever be taken for granted.
In summary, autopoiesis is the information—and associated abilities—that you need to have (repair, maintenance and differential reproduction) in order to keep the information that you want to have (e.g. vacuum cleaner functionality) alive and in good condition to ensure both your survival and that of your descendants. In a parallel way, my humanity is what I personally value, so my autopoietic capability is the repair, maintenance and differential reproductive capacity that I have to maintain my humanity and to share it with my descendants. The egg and sperm that produced me knew nothing of this, but the information was encoded there and only reached fruition six decades later as I sit here writing this—the inverse causality of autopoiesis.
Summary
There are three lines of reasoning pointing to the conclusion that autopoiesis provides a compelling case for the intelligent design of life.
• If life began in some stepwise manner from a non-autopoietic beginning, then autopoiesis will be the end product of some long and blind process of accidents and natural selection. Such a result would mean that autopoiesis is not essential to life, so some organisms should exist that never attained it, and some organisms should have lost it by natural selection because they do not need it. However, autopoiesis is universal in all forms of life, so it must be essential. The argument from the Second Law of Thermodynamics as applied to the vacuum cleaner analogy also points to the same conclusion. Both arguments demonstrate that autopoiesis is required at the beginning for life to even exist and perpetuate itself, and could not have turned up at the end of some long naturalistic process. This conclusion is consistent with the experimental finding that origin-of-life projects which begin without autopoiesis as a pre-requisite have proved universally futile in achieving even the first step towards life.
• Each level of the autopoietic hierarchy is dependent upon the one below it, but is causally separated from it by a Polanyi impossibility. Autopoiesis therefore cannot be reduced to any sequence of naturalistic causes.
• There is an unbridgeable abyss below the autopoietic hierarchy, between the dirty, mass-action chemistry of the natural environment and the perfect purity, the single-molecule precision, the structural specificity, and the inversely causal integration, regulation, repair, maintenance and differential reproduction of life.
TOPICS: News/Current Events
KEYWORDS: agenda; alexwilliams; bush; catholic; christian; congress; creation; deduve; dembski; dondeyoung; evolution; geraldschroeder; gerhart; globalwarming; guillermogonzalez; homosexualagenda; hubertyockey; intelligentdesign; jonathansarfati; kirschner; michaelbehe; michaelpolanyi; moralabsolutes; obama; peterborger; prolife; royaltruman; russellhumphreys; scotus; seanpitman; talkradio; williamdembski
Navigation: use the links below to view more comments.
first previous 1-20 ... 421-440, 441-460, 461-480 ... 901-918 next last
To: Coyoteman
There is so much anti-science nonsense spread about here its not worth even reading it. It isn't anti-science, it is emotion. Emotion rules here and at DU. All the usual suspects have a visceral gut reaction when their emotions are challenged (as do we all). I am just amazed at the lengths they will go to to quell the cognitive dissonance that they feel. It must be emotional hell to rationally know that what they believe in their heart is wrong.
441
posted on
01/12/2009 10:16:01 PM PST
by
LeGrande
(I once heard a smart man say that you canÂ’t reason someone out of something that they didnÂ’t reaso)
To: LeGrande
[[It must be emotional hell to rationally know that what they believe in their heart is wrong.]]
I weouldn’t know- All I know is that the science is for intelligent design, and against common descent as demonstrated many times here on FR.
To: Inappropriate Laughter
How many genetic nosepicking arguments are there for probabilities? Quite an umber it woudl appear. Yell me the story about hte moot improbabilites again- I just love that story.
1: It seems likely that RNA has the capability to support life based on RNA genomes that are copied and maintained through the catalytic function of RNA. There are substantial gaps, however, in scientific understanding concerning how the RNA world arose, the degree of metabolic complexity that it attained, and the way that it led to DNA genomes and protein enzymes.
2: Depending on the nature of the prebiotic environment, available building blocks may have included amino acids, hydroxy acids, sugars, purines, pyrimidines and fatty acids. These could have combined to form polymers of largely random sequence and mixed stereochemistry (handedness). Some of the polymers may have had special properties, such as adherence to a particular mineral surface, unusual resistance to degradation, or the propensity to form supramolecular aggregates. Eventually every polymer, no matter how stable, would have succumbed to degradation.
3: When the environment is altered, the special properties associated with a particular polymer may no longer apply and the capacity for self-replication may be lost. Persistence in a changing environment requires a more general mechanism for self-replication that allows the polymer sequence to change somewhat over time, but retain its heritage in most of the sequence that is unchanged.
4: If the building blocks of RNA were available in the prebiotic environment, if these combined to form polynucleotides, and if some of the polynucleotides began to self-replicate, then the RNA world may have emerged as the first form of life on Earth. But based on current knowledge of prebiotic chemistry, this is unlikely to have been the case. Ribose, phosphate, purines and pyrimidines all may have been available, although the case for pyrimidines is less compelling.
5: It is difficult to visualize a mechanism for self-replication that either would be impartial to these compositional differences or would treat them as sequence information in a broader sense and maintain them as heritable features.
6: The chief obstacle to understanding the origin of RNA-based life is identifying a plausible mechanism for overcoming the clutter wrought by prebiotic chemistry.
7: Another approach is to hypothesize that life did not begin with RNA; some other genetic system preceded RNA, just as it preceded DNA and protein. This approach has met with substantial progress in recent years, despite the lack of guidance from known metabolic pathways in biology regarding the chemical nature of a precursor to RNA.
8: It is also possible that RNA-based life was preceded by a replicating, evolving polymer that bore no resemblance to nucleic acids. Self-replication without darwinian evolution has been demonstrated for certain peptides and even small organic compounds. Why not cast the net broadly and consider any polymer that is capable of self-replication? A critical issue then becomes whether there is a sufficient diversity of polymer sequences that can be replicated faithfully to provide the basis for darwinian evolution.
9: The catalytic potential of TNA, PNA and other proposed precursors to RNA has not yet been explored, but any cogent hypothesis regarding pre-RNA life must consider whether that prior genetic system could have facilitated the appearance of RNA.
10: There is no known ribozyme in biology that catalyses the template-directed polymerization of NTPs [nucleoside 5’-triphosphates, a putative building block for RNA], but such molecules have been obtained using test-tube evolution. Like the evolution of organisms in nature, evolution of RNA in the laboratory involves repeated rounds of selective amplification, linking the survival of an RNA species to its fitness. In the laboratory, fitness is defined by the experimenter, for example, based on the ability of RNA to catalyse a particular chemical reaction.
11: A pool of one copy each of all possible 40mers [RNA chains of 40 nucleotides], with a mass of 26 kg, just might be achievable, but it is not clear if 40 nucleotides are sufficient to provide robust RNA polymerase activity. ... As a rule of thumb, the error rate of replication per nucleotide must be no more than about the inverse of genome length, corresponding to 99% fidelity for replication of a 100mer and 97.5% fidelity for replication of a 40mer. There may be polymerase ribozymes that meet these requirements, although such molecules have not yet been demonstrated.
12: The above discussion ignores other obstacles to RNA-catalysed RNA replication, such as maintaining a supply of activated mononucleotides, ensuring that the ribozyme will recognize its corresponding genomic RNA while ignoring other RNAs in the environment, overcoming stable self-structure within the template strand, separating the template and product strands, and operating in a similar manner on the product strand to generate new copies of the template. Additional genetic information might be required to overcome these obstacles, but a longer genome would necessitate an even higher fidelity of replication.
13: Although the central process of the RNA world was the replication of RNA genomes, some form of metabolism must have supported the process. In keeping with the second law of thermodynamics, the increase in order that occurs in a genetic system is achieved through the expenditure of high-energy starting materials that are converted to lower-energy products.
14: There are several important reactions in nucleotide synthesis that have not yet been carried out with a ribozyme. ... The possibility of a more complex RNA-based metabolism is purely conjectural.
15: Other RNA-based functions for which there is no evidence in biology, such as nucleotide synthesis and RNA polymerization, are assumed to have existed in the RNA world based on first principles, but it is important to recognize that this assumption is not supported by available historical evidence.
16: It is often said, again based on first principles rather than historical evidence, that RNA-based life must have entailed some form of cellular compartmentalization.
17: Although RNA is well suited as a genetic molecule and can evolve to perform a broad range of catalytic tasks, it has limited chemical functionality and thus may not be equipped to meet certain challenges and opportunities that arise in the environment. An important innovation of life on Earth was the development of a separate macromolecule that would be responsible for most catalytic functions, even though that molecule contained subunits that were poorly suited for replication. The invention of protein synthesis, instructed and catalysed by RNA, was the crowning achievement of the RNA world, but also began its demise.
18: It is not known whether the invention of protein synthesis preceded or followed the invention of DNA genomes. The primary advantage of DNA over RNA as a genetic material is the greater chemical stability of DNA, allowing much larger genomes based on DNA. Protein synthesis may require more genetic information than can be maintained by RNA.
19: A largely open question concerns the origin of the genetic code. The aminoacylation of RNA initially must have provided some selective advantage unrelated to the eventual development of a translation machinery.
20: The next step towards the origin of the genetic code was the formation of peptide bonds between amino acids that were attached to RNA. The products of this reaction must have conferred some selective advantage, even though the peptides probably would have been too small and too heterogeneous in sequence to function as catalysts. ... It is not clear, however, how the detailed assignments of the genetic code were made.
21: Insight into the origin and operation of the RNA world is largely inferential, based on the known chemical and biochemical properties of RNA. In the best of circumstances those inferences are supported by examining the role of RNA in contemporary biology. Without that support one must be careful not to draw detailed conclusions regarding these historical events.
“How many show-stoppers does it take to stop a show? Every one of Joyce’s listed problems (and there are others), is serious enough to forbid progress toward life. It’s a tale only an atheist could love, but here it is prominently featured in Nature. Notice that we are not picking on straw men, but the best champions the Darwin Party has to offer. Gerald Joyce, Mr. RNA World himself, lays out the latest and greatest story in the world’s most prestigious science journal. Are you impressed?”
http://www.creationsafaris.com/crev0702.htm
To: CottShop
[[On what basis do you assert that humans already had the metainfo necessary to understand language before there was any such thing as language?]]Reasonability-...
So you make up a scenario for which you have no evidence, and you expect me to accept it because it sounds "reasonable" to you? Frankly, it sounds "ludicrous" to me. Now what are we supposed to do--just grant that you're right because you're a better judge of what's reasonable than I am? This is what's supposed to pass for scientific discussion?
To: CottShop; GodGunsGuts
You’ve addressed NONE of the main central issues...William’s grasp of science is just fine- how bout getting back on topic? As I said before, I thought the topic here was Williams' paper. You seem to want to argue that his reasoning must be valid because he comes to the right conclusion. We were invited to examine what GGG considers an "airtight" argument. I've chosen to deal with that specific question, and my conclusion is that Williams' argument is logically flawed and based on false premises. There are plenty of other topics for arguing the "main central issues."
To: Ha Ha Thats Very Logical
[[So you make up a scenario for which you have no evidence, and you expect me to accept it because it sounds “reasonable” to you?]]
Yup- nwext question? Tell me Haha- have you got evidence that language evovled ever so slowly until the ‘metainfo’ of grammar was achieved? No? Then is your reasoning o nthe matter any more or less valid than mine? Meta info is obviously available concerning sounds, as someone hearing an alarming sound for hte first time will instinctivvely recoil- if you can show that metainfo such as this evolved slowly, then let’s have it. The argument that language supposedly came via syllable, word, then sentances then grammar isn’t an argument against metainfo of livign systems
Is the argument any less scientific than someone suggesting thast Macroevolution despite a complete lack of evidence, and myriad evidences against it’s biological, mathematical and natural possibilites happened anyways? No? And yet you fully accept that as ‘science’? Why? Because they’ve backed their claims up with evidence?
the question was whether metainfo in livign systems is capable of arising- you pointed out a non livign system and offered it as evidence? And that passes for a scientific discussion?
To: Ha Ha Thats Very Logical
[[As I said before, I thought the topic here was Williams’ paper. You seem to want to argue that his reasoning must be valid because he comes to the right conclusion. We were invited to examine what GGG considers an “airtight” argument. I’ve chosen to deal with that specific question, and my conclusion is that Williams’ argument is logically flawed and based on false premises. There are plenty of other topics for arguing the “main central issues.”]]
the one point does NOT invalidate the whole- the central issues- The discussion wasn’t about whether it’s airtight- the issue is whether or not naturalism can account for chemical purity, and whehter metainfo can arise naturally in living systems despite htere being a lack of info i nthe first place IF macroevolution were a possibility and htings evolved naturally.
you have chosen to ignore hte central issues- GGG made it clear that he was askign about these central claims and whether or not they could be refuted numerous times, but you have contented yourself and resigned yourself to apaprently accepting that hte paper must be invalid as a whoel because a few irrelevent points are not compeltely accurate evidently?
To: CottShop
Yup- nwext question? Tell me Haha- have you got evidence that language evovled ever so slowly until the ‘metainfo’ of grammar was achieved? People do study this stuff, you know. I recommend this book if you're really interested in learning how languages develop. Key passage for our purposes:
The evidence relied on in this book did not come from fossils or artifacts, but from the behavior of language itself, as observed in the present and the attested period [i.e., the period for which we have written records]....In other words, the underlying thesis is that the principles and processes of linguistic change in the distant past must have been similar to those that can be observed in action over the last 5,000 years or so.
There are elements of grammar common to all modern languages that are not seen, or are seen in only rudimentary forms, in the most ancient languages we have records for. So yes, there is evidence that grammar--the metainfo, if you will--continued to evolve after there were words. (Mind you, I don't expect you to accept this--the theory you just made up is obviously just as good as the ideas of people that spend their entire lives studying the history of language.)
the question was whether metainfo in livign systems is capable of arising- you pointed out a non livign system and offered it as evidence?
The comparison of language to living systems was Polanyi's. Remember Polanyi--the guy whose ideas are supposedly the basis for the paper we're discussing?
To: Ha Ha Thats Very Logical
(A) All aspects of life (not just bacterial flagellums and blood clotting cascades) lie beyond the reach of naturalistic explanations, and (B) only intelligent design meets the criterion of an acceptable historical inference according to the Law of Cause and Effect.
While in the other thread GGG did state he velt it might be an airtight case, this thread is abotu htese two central issues
[[Polanyi argued that living organisms have a machine-like structure that cannot be explained by (or reduced to) the physics and chemistry of the molecules of which they consist.]]
What is beign discussed in htis htread is whether “All of these links are also ‘choreographed’ by information—a phenomenon that never occurs in the natural environment.” You have tried to take the discussion from living systems to non living systems, and hten claim it pokes holes in the argument, just as Polanyi claimed Grammar poked holes in the idea that livign metainfo couldn’t arise naturally. The individual steps of language however are not the metainfo we’re discussing in livign systems, however, metainfo can deal with any one of htose steps to help preserve the species IF those steps include an advantage for the living system/species- it’s late- I’m tired, will think this htrough more tomorrow with a fresher mind
To: CottShop; GodGunsGuts
The discussion wasn’t about whether it’s airtight GGG in
another topic: "Actually, I think I have stumbled onto a creationist paper that makes an airtight case that materialist evolution is impossible. Anyone care to read the paper and take me up on my challenge?"
To: CottShop
You rendered my reply irrelevant by 14 seconds! But you’re right about one thing—it’s late. See you tomorrow.
To: Ha Ha Thats Very Logical
[[ So yes, there is evidence that grammar—the metainfo, if you will—continued to evolve after there were words.]]
Again, you are tryign to equate metainfo in living systems with simplistic non livign ‘metainfo’ of non living systems of comunication.
[[The comparison of language to living systems was Polanyi’s.]]
and you are furthering the artument apparently htinking it pokes holes in the argument that living sdystem’s metainfo could not have arisen naturally.
[[(Mind you, I don’t expect you to accept this—the theory you just made up is obviously just as good as the ideas of people that spend their entire lives studying the history of language.)]]
No need to get nasty- My point is that the metainfo is NOT the language, as claimed by polanyi- it may be ‘metainfo’ in regards to the non living system of language- but it is magnititudes less complex than living systems inherrent metainfo and irrelevent to the discussion and question about whehter living system metainfo can arise naturally.
To: Ha Ha Thats Very Logical
I know hwat he said i nthe other thread- however he made his itnentions for htis thread quite clear several times in both these threads
(A) All aspects of life (not just bacterial flagellums and blood clotting cascades) lie beyond the reach of naturalistic explanations, and (B) only intelligent design meets the criterion of an acceptable historical inference according to the Law of Cause and Effect.
good night.
453
posted on
01/12/2009 11:37:26 PM PST
by
CottShop
(uite)
To: GodGunsGuts
What does this have to do with anything under the keyword homosexualagenda?
454
posted on
01/12/2009 11:46:26 PM PST
by
NinoFan
To: NinoFan
Anyone can add a keyword. Half the listed keywords on this thread are irrelevant.
455
posted on
01/13/2009 2:03:45 AM PST
by
shibumi
(...so if it's organic, where are its organs?)
To: shibumi
That’s true. The mods used to police that a bit better though. Someone could be called out for ‘keyword abuse’.
456
posted on
01/13/2009 2:07:49 AM PST
by
NinoFan
To: CottShop
That is correct- it is over whehter it is sound or not- IF it is, then yes, I’d agree debate for macroevolution has taken a serious blow once again- IF it is not, then all macroevolution has to deal with then are hte other myriad of serious blows.I believe we established that the theory disallows abiogenesis, but not evolution. If the theory is sound, then life cannot arise from natural processes, it must be designed. Once designed and created, there is nothing in the theory that disallows a design that permits the organism to evolve. Macroevolution will have to wait for another day.
457
posted on
01/13/2009 3:46:18 AM PST
by
tacticalogic
("Oh bother!" said Pooh, as he chambered his last round.)
To: Coyoteman
The lessons of Dover are two:
(1) Local School Boards are not a good vehicle for a long legal fight, because the electoral turnover is too quick -- as quick as a whim.
(2) Judges make science. In today's culture science is dicta. That is what you wanted, right?
458
posted on
01/13/2009 4:47:26 AM PST
by
bvw
To: LeGrande
I’d can understand people’s confusions with your mighty words. Maybe you should provide diagrams.
459
posted on
01/13/2009 4:49:37 AM PST
by
bvw
To: LeGrande
M=F/A So you are saying that if something is at rest it has zero mass too? No, I am not saying anything of the sort. Those are your ideas. M=F/A, if A=0, M is undefined. Basic algebra dictates that there can not be a zero in the denominator.
Navigation: use the links below to view more comments.
first previous 1-20 ... 421-440, 441-460, 461-480 ... 901-918 next last
Disclaimer:
Opinions posted on Free Republic are those of the individual
posters and do not necessarily represent the opinion of Free Republic or its
management. All materials posted herein are protected by copyright law and the
exemption for fair use of copyrighted works.
FreeRepublic.com is powered by software copyright 2000-2008 John Robinson