Skip to comments.
Scientists find missing link between whale and its closest relative, the hippo
UC Berkeley News ^
| 24 January 2005
| Robert Sanders, Media Relations
Posted on 02/08/2005 3:50:43 AM PST by PatrickHenry
A group of four-footed mammals that flourished worldwide for 40 million years and then died out in the ice ages is the missing link between the whale and its not-so-obvious nearest relative, the hippopotamus.
The conclusion by University of California, Berkeley, post-doctoral fellow Jean-Renaud Boisserie and his French colleagues finally puts to rest the long-standing notion that the hippo is actually related to the pig or to its close relative, the South American peccary. In doing so, the finding reconciles the fossil record with the 20-year-old claim that molecular evidence points to the whale as the closest relative of the hippo.
"The problem with hippos is, if you look at the general shape of the animal it could be related to horses, as the ancient Greeks thought, or pigs, as modern scientists thought, while molecular phylogeny shows a close relationship with whales," said Boisserie. "But cetaceans – whales, porpoises and dolphins – don't look anything like hippos. There is a 40-million-year gap between fossils of early cetaceans and early hippos."
In a paper appearing this week in the Online Early Edition of the Proceedings of the National Academy of Sciences, Boisserie and colleagues Michel Brunet and Fabrice Lihoreau fill in this gap by proposing that whales and hippos had a common water-loving ancestor 50 to 60 million years ago that evolved and split into two groups: the early cetaceans, which eventually spurned land altogether and became totally aquatic; and a large and diverse group of four-legged beasts called anthracotheres. The pig-like anthracotheres, which blossomed over a 40-million-year period into at least 37 distinct genera on all continents except Oceania and South America, died out less than 2 and a half million years ago, leaving only one descendent: the hippopotamus.
This proposal places whales squarely within the large group of cloven-hoofed mammals (even-toed ungulates) known collectively as the Artiodactyla – the group that includes cows, pigs, sheep, antelopes, camels, giraffes and most of the large land animals. Rather than separating whales from the rest of the mammals, the new study supports a 1997 proposal to place the legless whales and dolphins together with the cloven-hoofed mammals in a group named Cetartiodactyla.
"Our study shows that these groups are not as unrelated as thought by morphologists," Boisserie said, referring to scientists who classify organisms based on their physical characteristics or morphology. "Cetaceans are artiodactyls, but very derived artiodactyls."
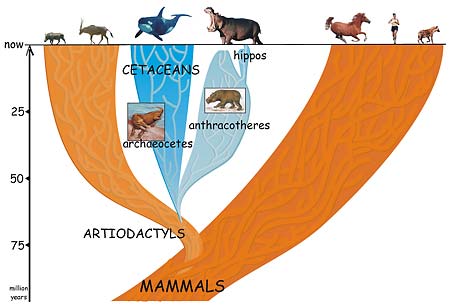
The origin of hippos has been debated vociferously for nearly 200 years, ever since the animals were rediscovered by pioneering French paleontologist Georges Cuvier and others. Their conclusion that hippos are closely related to pigs and peccaries was based primarily on their interpretation of the ridges on the molars of these species, Boisserie said.
"In this particular case, you can't really rely on the dentition, however," Boisserie said. "Teeth are the best preserved and most numerous fossils, and analysis of teeth is very important in paleontology, but they are subject to lots of environmental processes and can quickly adapt to the outside world. So, most characteristics are not dependable indications of relationships between major groups of mammals. Teeth are not as reliable as people thought."
As scientists found more fossils of early hippos and anthracotheres, a competing hypothesis roiled the waters: that hippos are descendents of the anthracotheres.
All this was thrown into disarray in 1985 when UC Berkeley's Vincent Sarich, a pioneer of the field of molecular evolution and now a professor emeritus of anthropology, analyzed blood proteins and saw a close relationship between hippos and whales. A subsequent analysis of mitochondrial, nuclear and ribosomal DNA only solidified this relationship.
Though most biologists now agree that whales and hippos are first cousins, they continue to clash over how whales and hippos are related, and where they belong within the even-toed ungulates, the artiodactyls. A major roadblock to linking whales with hippos was the lack of any fossils that appeared intermediate between the two. In fact, it was a bit embarrassing for paleontologists because the claimed link between the two would mean that one of the major radiations of mammals – the one that led to cetaceans, which represent the most successful re-adaptation to life in water – had an origin deeply nested within the artiodactyls, and that morphologists had failed to recognize it.
This new analysis finally brings the fossil evidence into accord with the molecular data, showing that whales and hippos indeed are one another's closest relatives.
"This work provides another important step for the reconciliation between molecular- and morphology-based phylogenies, and indicates new tracks for research on emergence of cetaceans," Boisserie said.
Boisserie became a hippo specialist while digging with Brunet for early human ancestors in the African republic of Chad. Most hominid fossils earlier than about 2 million years ago are found in association with hippo fossils, implying that they lived in the same biotopes and that hippos later became a source of food for our distant ancestors. Hippos first developed in Africa 16 million years ago and exploded in number around 8 million years ago, Boisserie said.
Now a post-doctoral fellow in the Human Evolution Research Center run by integrative biology professor Tim White at UC Berkeley, Boisserie decided to attempt a resolution of the conflict between the molecular data and the fossil record. New whale fossils discovered in Pakistan in 2001, some of which have limb characteristics similar to artiodactyls, drew a more certain link between whales and artiodactyls. Boisserie and his colleagues conducted a phylogenetic analysis of new and previous hippo, whale and anthracothere fossils and were able to argue persuasively that anthracotheres are the missing link between hippos and cetaceans.
While the common ancestor of cetaceans and anthracotheres probably wasn't fully aquatic, it likely lived around water, he said. And while many anthracotheres appear to have been adapted to life in water, all of the youngest fossils of anthracotheres, hippos and cetaceans are aquatic or semi-aquatic.
"Our study is the most complete to date, including lots of different taxa and a lot of new characteristics," Boisserie said. "Our results are very robust and a good alternative to our findings is still to be formulated."
Brunet is associated with the Laboratoire de Géobiologie, Biochronologie et Paléontologie Humaine at the Université de Poitiers and with the Collège de France in Paris. Lihoreau is a post-doctoral fellow in the Département de Paléontologie of the Université de N'Djaména in Chad.
The work was supported in part by the Mission Paléoanthropologique Franco-Tchadienne, which is co-directed by Brunet and Patrick Vignaud of the Université de Poitiers, and in part by funds to Boisserie from the Fondation Fyssen, the French Ministère des Affaires Etrangères and the National Science Foundation's Revealing Hominid Origins Initiative, which is co-directed by Tim White and Clark Howell of UC Berkeley.
TOPICS: Culture/Society; Miscellaneous; Philosophy
KEYWORDS: crevolist; darwin; evolution; whale
Navigation: use the links below to view more comments.
first previous 1-20 ... 1,901-1,920, 1,921-1,940, 1,941-1,960 ... 2,241-2,242 next last
To: Tribune7
You wrote:
But the creationist argument has as much weight as yours... Then I wrote: I do not believe it does. What empirical evidence supports creationism? What predictions does creationism make about biology? How would we test any predictions made by creationism?
You replied: Hey, I don't have to be convinced that evolution provides accurate answers to much about nature -- just not as many as its proponents claim, and, especially, not to the point where it can't be questioned.
Then I said: Theories, including evolution, were meant to be questioned. Is creationism?
To which you replied: Do mean the existence of God or a particular intpretation of Genesis?
I meant creationism.
Is creationism meant to be tested? How does one formulate a test of creationist theory? Scientific theories are designed to validated, or invalidated, through experimentation. How does one go about designing a test to validate creationism?
1,921
posted on
02/11/2005 1:26:10 PM PST
by
Liberal Classic
(No better friend, no worse enemy. Semper Fi.)
To: King Prout
one of my personal favorites.Indeed! It's a good one. One of the interesting parts was when G3K responded to all five supposed predictions and not a single evo responded:
1. There should be no transitional species.
There are not any. What transitional species is there between reptiles and mammals? What evidence is there for it? As often pointed out it is in the greatest changes that there should be the most evidence. For this the greatest change in the supposed evolutionary tree of life there is absolutely no evidence.
2. There are most certainly no pre-human species.
The origin of humans - homo sapiens - has never been determined in any credible way. The closest species to homo sapiens in chronology - Neanderthals - has been proven not to have been a possible ancestor of man.
BTW - the statement is one of the rhetorical nonsenses used by evolutionists. Bacteria are pre-human.
3. There should be no evidence, whether in fossils or DNA, showing the chronological evolution of life.
There is not. There are numerous fossils and numerous gaps in the fossil record. Species appear fully formed in numerous places with absolutely no ancestry. The Cambrian explosion is a complete refutation of evolution and led Gould and other evolutionists to break with Darwinian evolution.
4. There must surely be at least one species, and probably several, having no genetic similarities with any other life on earth.
There is no such requirement in creationism. In fact, that all life is based on DNA is abundant proof that abiogenesis (life from non-life) is impossible. Thus the materialistic basis of evolutionism is totally discredited since it leaves as the only possible source of life the intervention of a Creator.
5. The fossil record must show all kinds of species (such as dinosaurs and humans) living together at the same time.
Nonsense. Another total non-prediction of creationism. Evolution proposes constant changes in environment and constant mutations as the source of the transformation of species. That many species have remained unchanged for hundreds of millions of years without change shows the evolutionist predictions to be false.
1,922
posted on
02/11/2005 1:26:26 PM PST
by
Michael_Michaelangelo
(The best theory is not ipso facto a good theory. Lots of links on my homepage...)
To: Pantera
Hal is not an idiot. He is correct. The sun eliminates the claim of entropy through 2nd Law.
Anyone who claims the 2nd law invalidates evolution is just plain silly.
1,923
posted on
02/11/2005 1:28:51 PM PST
by
shubi
(Peace through superior firepower.)
To: Tribune7
Hal is obviously not a native English speaker. You should ask him to clarify something you don't understand, instead of flaming him.
He knows his science pretty well. YOu might be able to learn from him and he might be converted to literalist fundamentalism by you. ;-)
1,924
posted on
02/11/2005 1:32:37 PM PST
by
shubi
(Peace through superior firepower.)
To: Tribune7
Don't you understand the difference between mountains of evidence (evolution) and vacuous arguments (creationism)?
1,925
posted on
02/11/2005 1:34:05 PM PST
by
shubi
(Peace through superior firepower.)
To: Tribune7
Evolution can only be questioned if you have some evidence that refutes it. What evidence do you have?
1,926
posted on
02/11/2005 1:36:15 PM PST
by
shubi
(Peace through superior firepower.)
1,927
posted on
02/11/2005 1:42:03 PM PST
by
Michael_Michaelangelo
(The best theory is not ipso facto a good theory. Lots of links on my homepage...)
From the link:
Evolutionists almost since the start have tried to silence opponents. While they constantly claim to be scientists, it seems that instead of following the principles of science - questioning, discussion, and challenging of existing theories, they follow the principles of ideology - silencing and destroying opponents.
Does this describe you? If so, you may not be a scientist.
1,928
posted on
02/11/2005 1:43:44 PM PST
by
Michael_Michaelangelo
(The best theory is not ipso facto a good theory. Lots of links on my homepage...)
To: shubi
shibu, I couldn't care less about the evolution versus creationism controversy.
The evidence clearly supports many aspects of your AGENDA and science may eventually have sufficient evidence to support evolution from one species to another. Right now we have only determined and witnessed cross breeding within a particular species. I believe that eventually we will have a much greater amount of data to work from and yes, it is highly probable that is will support the whole evolutionary picture. But until then my questions are valid and your propaganda is based on lies.
To: Liberal Classic
You wrote: But the creationist argument has as much weight as yours... You have to take into account that was a response to the specific claim Why would an "intelligent designer" make the same design error in only two lines of heredity and not in all of them?
And of course the creationist argument has as much weight as that.
Now, the existance of a Creator is not meant to be tested but should be considered culturally axiomatic unless you want to scrap the Declaration of Independence. Further, science and reason far more strongly indicate the existence of a Creator than not.
What can and should be tested is any claim as to how the Creator did something.
To: Michael_Michaelangelo
Pred-There should be no transitional species.
Ans: There are not any.
Transition from synapsid reptiles to mammals
This is the best-documented transition between vertebrate classes. So far this series is known only as a series of genera or families; the transitions from species to species are not known. But the family sequence is quite complete. Each group is clearly related to both the group that came before, and the group that came after, and yet the sequence is so long that the fossils at the end are astoundingly different from those at the beginning. As Rowe recently said about this transition (in Szalay et al., 1993), "When sampling artifact is removed and all available character data analyzed [with computer phylogeny programs that do not assume anything about evolution], a highly corroborated, stable phylogeny remains, which is largely consistent with the temporal distributions of taxa recorded in the fossil record." Similarly, Gingerich has stated (1977) "While living mammals are well separated from other groups of animals today, the fossil record clearly shows their origin from a reptilian stock and permits one to trace the origin and radiation of mammals in considerable detail." For more details, see Kermack's superb and readable little book (1984), Kemp's more detailed but older book (1982), and read Szalay et al.'s recent collection of review articles (1993, vol. 1).
This list starts with pelycosaurs (early synapsid reptiles) and continues with therapsids and cynodonts up to the first unarguable "mammal". Most of the changes in this transition involved elaborate repackaging of an expanded brain and special sense organs, remodeling of the jaws & teeth for more efficient eating, and changes in the limbs & vertebrae related to active, legs-under-the-body locomotion. Here are some differences to keep an eye on:
# Early Reptiles Mammals
1 No fenestrae in skull Massive fenestra exposes all of braincase
2 Braincase attached loosely Braincase attached firmly to skull
3 No secondary palate Complete bony secondary palate
4 Undifferentiated dentition Incisors, canines, premolars, molars
5 Cheek teeth uncrowned points Cheek teeth (PM & M) crowned & cusped
6 Teeth replaced continuously Teeth replaced once at most
7 Teeth with single root Molars double-rooted
8 Jaw joint quadrate-articular Jaw joint dentary-squamosal (*)
9 Lower jaw of several bones Lower jaw of dentary bone only
10 Single ear bone (stapes) Three ear bones (stapes, incus, malleus)
11 Joined external nares Separate external nares
12 Single occipital condyle Double occipital condyle
13 Long cervical ribs Cervical ribs tiny, fused to vertebrae
14 Lumbar region with ribs Lumbar region rib-free
15 No diaphragm Diaphragm
16 Limbs sprawled out from body Limbs under body
17 Scapula simple Scapula with big spine for muscles
18 Pelvic bones unfused Pelvis fused
19 Two sacral (hip) vertebrae Three or more sacral vertebrae
20 Toe bone #'s 2-3-4-5-4 Toe bones 2-3-3-3-3
21 Body temperature variable Body temperature constant
(*) The presence of a dentary-squamosal jaw joint has been arbitrarily selected as the defining trait of a mammal.
Paleothyris (early Pennsylvanian) -- An early captorhinomorph reptile, with no temporal fenestrae at all.
Protoclepsydrops haplous (early Pennsylvanian) -- The earliest known synapsid reptile. Little temporal fenestra, with all surrounding bones intact. Fragmentary. Had amphibian-type vertebrae with tiny neural processes. (reptiles had only just separated from the amphibians)
Clepsydrops (early Pennsylvanian) -- The second earliest known synapsid. These early, very primitive synapsids are a primitive group of pelycosaurs collectively called "ophiacodonts".
Archaeothyris (early-mid Pennsylvanian) -- A slightly later ophiacodont. Small temporal fenestra, now with some reduced bones (supratemporal). Braincase still just loosely attached to skull. Slight hint of different tooth types. Still has some extremely primitive, amphibian/captorhinid features in the jaw, foot, and skull. Limbs, posture, etc. typically reptilian, though the ilium (major hip bone) was slightly enlarged.
Varanops (early Permian) -- Temporal fenestra further enlarged. Braincase floor shows first mammalian tendencies & first signs of stronger attachment to rest of skull (occiput more strongly attached). Lower jaw shows first changes in jaw musculature (slight coronoid eminence). Body narrower, deeper: vertebral column more strongly constructed. Ilium further enlarged, lower-limb musculature starts to change (prominent fourth trochanter on femur). This animal was more mobile and active. Too late to be a true ancestor, and must be a "cousin".
Haptodus (late Pennsylvanian) -- One of the first known sphenacodonts, showing the initiation of sphenacodont features while retaining many primitive features of the ophiacodonts. Occiput still more strongly attached to the braincase. Teeth become size-differentiated, with biggest teeth in canine region and fewer teeth overall. Stronger jaw muscles. Vertebrae parts & joints more mammalian. Neural spines on vertebrae longer. Hip strengthened by fusing to three sacral vertebrae instead of just two. Limbs very well developed.
Dimetrodon, Sphenacodon or a similar sphenacodont (late Pennsylvanian to early Permian, 270 Ma) -- More advanced pelycosaurs, clearly closely related to the first therapsids (next). Dimetrodon is almost definitely a "cousin" and not a direct ancestor, but as it is known from very complete fossils, it's a good model for sphenacodont anatomy. Medium-sized fenestra. Teeth further differentiated, with small incisors, two huge deep- rooted upper canines on each side, followed by smaller cheek teeth, all replaced continuously. Fully reptilian jaw hinge. Lower jaw bone made of multiple bones & with first signs of a bony prong later involved in the eardrum, but there was no eardrum yet, so these reptiles could only hear ground-borne vibrations (they did have a reptilian middle ear). Vertebrae had still longer neural spines (spectacularly so in Dimetrodon, which had a sail), and longer transverse spines for stronger locomotion muscles.
Biarmosuchia (late Permian) -- A therocephalian -- one of the earliest, most primitive therapsids. Several primitive, sphenacodontid features retained: jaw muscles inside the skull, platelike occiput, palatal teeth. New features: Temporal fenestra further enlarged, occupying virtually all of the cheek, with the supratemporal bone completely gone. Occipital plate slanted slightly backwards rather than forwards as in pelycosaurs, and attached still more strongly to the braincase. Upper jaw bone (maxillary) expanded to separate lacrymal from nasal bones, intermediate between early reptiles and later mammals. Still no secondary palate, but the vomer bones of the palate developed a backward extension below the palatine bones. This is the first step toward a secondary palate, and with exactly the same pattern seen in cynodonts. Canine teeth larger, dominating the dentition. Variable tooth replacement: some therocephalians (e.g Scylacosaurus) had just one canine, like mammals, and stopped replacing the canine after reaching adult size. Jaw hinge more mammalian in position and shape, jaw musculature stronger (especially the mammalian jaw muscle). The amphibian-like hinged upper jaw finally became immovable. Vertebrae still sphenacodontid-like. Radical alteration in the method of locomotion, with a much more mobile forelimb, more upright hindlimb, & more mammalian femur & pelvis. Primitive sphenacodontid humerus. The toes were approaching equal length, as in mammals, with #toe bones varying from reptilian to mammalian. The neck & tail vertebrae became distinctly different from trunk vertebrae. Probably had an eardrum in the lower jaw, by the jaw hinge.
Procynosuchus (latest Permian) -- The first known cynodont -- a famous group of very mammal-like therapsid reptiles, sometimes considered to be the first mammals. Probably arose from the therocephalians, judging from the distinctive secondary palate and numerous other skull characters. Enormous temporal fossae for very strong jaw muscles, formed by just one of the reptilian jaw muscles, which has now become the mammalian masseter. The large fossae is now bounded only by the thin zygomatic arch (cheekbone to you & me). Secondary palate now composed mainly of palatine bones (mammalian), rather than vomers and maxilla as in older forms; it's still only a partial bony palate (completed in life with soft tissue). Lower incisor teeth was reduced to four (per side), instead of the previous six (early mammals had three). Dentary now is 3/4 of lower jaw; the other bones are now a small complex near the jaw hinge. Jaw hinge still reptilian. Vertebral column starts to look mammalian: first two vertebrae modified for head movements, and lumbar vertebrae start to lose ribs, the first sign of functional division into thoracic and lumbar regions. Scapula beginning to change shape. Further enlargement of the ilium and reduction of the pubis in the hip. A diaphragm may have been present.
Dvinia [also "Permocynodon"] (latest Permian) -- Another early cynodont. First signs of teeth that are more than simple stabbing points -- cheek teeth develop a tiny cusp. The temporal fenestra increased still further. Various changes in the floor of the braincase; enlarged brain. The dentary bone was now the major bone of the lower jaw. The other jaw bones that had been present in early reptiles were reduced to a complex of smaller bones near the jaw hinge. Single occipital condyle splitting into two surfaces. The postcranial skeleton of Dvinia is virtually unknown and it is not therefore certain whether the typical features found at the next level had already evolved by this one. Metabolic rate was probably increased, at least approaching homeothermy.
Thrinaxodon (early Triassic) -- A more advanced "galesaurid" cynodont. Further development of several of the cynodont features seen already. Temporal fenestra still larger, larger jaw muscle attachments. Bony secondary palate almost complete. Functional division of teeth: incisors (four uppers and three lowers), canines, and then 7-9 cheek teeth with cusps for chewing. The cheek teeth were all alike, though (no premolars & molars), did not occlude together, were all single- rooted, and were replaced throughout life in alternate waves. Dentary still larger, with the little quadrate and articular bones were loosely attached. The stapes now touched the inner side of the quadrate. First sign of the mammalian jaw hinge, a ligamentous connection between the lower jaw and the squamosal bone of the skull. The occipital condyle is now two slightly separated surfaces, though not separated as far as the mammalian double condyles. Vertebral connections more mammalian, and lumbar ribs reduced. Scapula shows development of a new mammalian shoulder muscle. Ilium increased again, and all four legs fully upright, not sprawling. Tail short, as is necessary for agile quadrupedal locomotion. The whole locomotion was more agile. Number of toe bones is 2.3.4.4.3, intermediate between reptile number (2.3.4.5.4) and mammalian (2.3.3.3.3), and the "extra" toe bones were tiny. Nearly complete skeletons of these animals have been found curled up - a possible reaction to conserve heat, indicating possible endothermy? Adults and juveniles have been found together, possibly a sign of parental care. The specialization of the lumbar area (e.g. reduction of ribs) is indicative of the presence of a diaphragm, needed for higher O2 intake and homeothermy. NOTE on hearing: The eardrum had developed in the only place available for it -- the lower jaw, right near the jaw hinge, supported by a wide prong (reflected lamina) of the angular bone. These animals could now hear airborne sound, transmitted through the eardrum to two small lower jaw bones, the articular and the quadrate, which contacted the stapes in the skull, which contacted the cochlea. Rather a roundabout system and sensitive to low-frequency sound only, but better than no eardrum at all! Cynodonts developed quite loose quadrates and articulars that could vibrate freely for sound transmittal while still functioning as a jaw joint, strengthened by the mammalian jaw joint right next to it. All early mammals from the Lower Jurassic have this low-frequency ear and a double jaw joint. By the middle Jurassic, mammals lost the reptilian joint (though it still occurs briefly in embryos) and the two bones moved into the nearby middle ear, became smaller, and became much more sensitive to high-frequency sounds.
Cynognathus (early Triassic, 240 Ma; suspected to have existed even earlier) -- We're now at advanced cynodont level. Temporal fenestra larger. Teeth differentiating further; cheek teeth with cusps met in true occlusion for slicing up food, rate of replacement reduced, with mammalian-style tooth roots (though single roots). Dentary still larger, forming 90% of the muscle-bearing part of the lower jaw. TWO JAW JOINTS in place, mammalian and reptilian: A new bony jaw joint existed between the squamosal (skull) and the surangular bone (lower jaw), while the other jaw joint bones were reduced to a compound rod lying in a trough in the dentary, close to the middle ear. Ribs more mammalian. Scapula halfway to the mammalian condition. Limbs were held under body. There is possible evidence for fur in fossil pawprints.
Diademodon (early Triassic, 240 Ma; same strata as Cynognathus) -- Temporal fenestra larger still, for still stronger jaw muscles. True bony secondary palate formed exactly as in mammals, but didn't extend quite as far back. Turbinate bones possibly present in the nose (warm-blooded?). Dental changes continue: rate of tooth replacement had decreased, cheek teeth have better cusps & consistent wear facets (better occlusion). Lower jaw almost entirely dentary, with tiny articular at the hinge. Still a double jaw joint. Ribs shorten suddenly in lumbar region, probably improving diaphragm function & locomotion. Mammalian toe bones (2.3.3.3.3), with closely related species still showing variable numbers.
Probelesodon (mid-Triassic; South America) -- Fenestra very large, still separate from eyesocket (with postorbital bar). Secondary palate longer, but still not complete. Teeth double-rooted, as in mammals. Nares separated. Second jaw joint stronger. Lumbar ribs totally lost; thoracic ribs more mammalian, vertebral connections very mammalian. Hip & femur more mammalian.
Probainognathus (mid-Triassic, 239-235 Ma, Argentina) -- Larger brain with various skull changes: pineal foramen ("third eye") closes, fusion of some skull plates. Cheekbone slender, low down on the side of the eye socket. Postorbital bar still there. Additional cusps on cheek teeth. Still two jaw joints. Still had cervical ribs & lumbar ribs, but they were very short. Reptilian "costal plates" on thoracic ribs mostly lost. Mammalian #toe bones.
Exaeretodon (mid-late Triassic, 239Ma, South America) -- (Formerly lumped with the herbivorous gomphodont cynodonts.) Mammalian jaw prong forms, related to eardrum support. Three incisors only (mammalian). Costal plates completely lost. More mammalian hip related to having limbs under the body. Possibly the first steps toward coupling of locomotion & breathing. This is probably a "cousin" fossil not directly ancestral, as it has several new but non-mammalian teeth traits.
GAP of about 30 my in the late Triassic, from about 239-208 Ma. Only one early mammal fossil is known from this time. The next time fossils are found in any abundance, tritylodontids and trithelodontids had already appeared, leading to some very heated controversy about their relative placement in the chain to mammals. Recent discoveries seem to show trithelodontids to be more mammal- like, with tritylodontids possibly being an offshoot group (see Hopson 1991, Rowe 1988, Wible 1991, and Shubin et al. 1991). Bear in mind that both these groups were almost fully mammalian in every feature, lacking only the final changes in the jaw joint and middle ear.
Oligokyphus, Kayentatherium (early Jurassic, 208 Ma) -- These are tritylodontids, an advanced cynodont group. Face more mammalian, with changes around eyesocket and cheekbone. Full bony secondary palate. Alternate tooth replacement with double-rooted cheek teeth, but without mammalian-style tooth occlusion (which some earlier cynodonts already had). Skeleton strikingly like egg- laying mammals (monotremes). Double jaw joint. More flexible neck, with mammalian atlas & axis and double occipital condyle. Tail vertebrae simpler, like mammals. Scapula is now substantially mammalian, and the forelimb is carried directly under the body. Various changes in the pelvis bones and hind limb muscles; this animal's limb musculature and locomotion were virtually fully mammalian. Probably cousin fossils (?), with Oligokyphus being more primitive than Kayentatherium. Thought to have diverged from the trithelodontids during that gap in the late Triassic. There is disagreement about whether the tritylodontids were ancestral to mammals (presumably during the late Triassic gap) or whether they are a specialized offshoot group not directly ancestral to mammals.
Pachygenelus, Diarthrognathus (earliest Jurassic, 209 Ma) -- These are trithelodontids, a slightly different advanced cynodont group. New discoveries (Shubin et al., 1991) show that these animals are very close to the ancestry of mammals. Inflation of nasal cavity, establishment of Eustachian tubes between ear and pharynx, loss of postorbital bar. Alternate replacement of mostly single- rooted teeth. This group also began to develop double tooth roots -- in Pachygenelus the single root of the cheek teeth begins to split in two at the base. Pachygenelus also has mammalian tooth enamel, and mammalian tooth occlusion. Double jaw joint, with the second joint now a dentary-squamosal (instead of surangular), fully mammalian. Incipient dentary condyle. Reptilian jaw joint still present but functioning almost entirely in hearing; postdentary bones further reduced to tiny rod of bones in jaw near middle ear; probably could hear high frequencies now. More mammalian neck vertebrae for a flexible neck. Hip more mammalian, with a very mammalian iliac blade & femur. Highly mobile, mammalian-style shoulder. Probably had coupled locomotion & breathing. These are probably "cousin" fossils, not directly ancestral (the true ancestor is thought to have occurred during that late Triassic gap). Pachygenelus is pretty close, though.
Adelobasileus cromptoni (late Triassic; 225 Ma, west Texas) -- A recently discovered fossil proto-mammal from right in the middle of that late Triassic gap! Currently the oldest known "mammal." Only the skull was found. "Some cranial features of Adelobasileus, such as the incipient promontorium housing the cochlea, represent an intermediate stage of the character transformation from non-mammalian cynodonts to Liassic mammals" (Lucas & Luo, 1993). This fossil was found from a band of strata in the western U.S. that had not previously been studied for early mammals. Also note that this fossil dates from slightly before the known tritylodonts and trithelodonts, though it has long been suspected that tritilodonts and trithelodonts were already around by then. Adelobasileus is thought to have split off from either a trityl. or a trithel., and is either identical to or closely related to the common ancestor of all mammals.
Sinoconodon (early Jurassic, 208 Ma) -- The next known very ancient proto-mammal. Eyesocket fully mammalian now (closed medial wall). Hindbrain expanded. Permanent cheekteeth, like mammals, but the other teeth were still replaced several times. Mammalian jaw joint stronger, with large dentary condyle fitting into a distinct fossa on the squamosal. This final refinement of the joint automatically makes this animal a true "mammal". Reptilian jaw joint still present, though tiny.
Kuehneotherium (early Jurassic, about 205 Ma) -- A slightly later proto-mammal, sometimes considered the first known pantothere (primitive placental-type mammal). Teeth and skull like a placental mammal. The three major cusps on the upper & lower molars were rotated to form interlocking shearing triangles as in the more advanced placental mammals & marsupials. Still has a double jaw joint, though.
Eozostrodon, Morganucodon, Haldanodon (early Jurassic, ~205 Ma) -- A group of early proto-mammals called "morganucodonts". The restructuring of the secondary palate and the floor of the braincase had continued, and was now very mammalian. Truly mammalian teeth: the cheek teeth were finally differentiated into simple premolars and more complex molars, and teeth were replaced only once. Triangular- cusped molars. Reversal of the previous trend toward reduced incisors, with lower incisors increasing to four. Tiny remnant of the reptilian jaw joint. Once thought to be ancestral to monotremes only, but now thought to be ancestral to all three groups of modern mammals -- monotremes, marsupials, and placentals.
Peramus (late Jurassic, about 155 Ma) -- A "eupantothere" (more advanced placental-type mammal). The closest known relative of the placentals & marsupials. Triconodont molar has with more defined cusps. This fossil is known only from teeth, but judging from closely related eupantotheres (e.g. Amphitherium) it had finally lost the reptilian jaw joint, attaing a fully mammalian three-boned middle ear with excellent high-frequency hearing. Has only 8 cheek teeth, less than other eupantotheres and close to the 7 of the first placental mammals. Also has a large talonid on its "tribosphenic" molars, almost as large as that of the first placentals -- the first development of grinding capability.
Endotherium (very latest Jurassic, 147 Ma) -- An advanced eupantothere. Fully tribosphenic molars with a well- developed talonid. Known only from one specimen. From Asia; recent fossil finds in Asia suggest that the tribosphenic molar evolved there.
Kielantherium and Aegialodon (early Cretaceous) -- More advanced eupantotheres known only from teeth. Kielantherium is from Asia and is known from slightly older strata than the European Aegialodon. Both have the talonid on the lower molars. The wear on it indicates that a major new cusp, the protocone, had evolved on the upper molars. By the Middle Cretaceous, animals with the new tribosphenic molar had spread into North America too (North America was still connected to Europe.)
Steropodon galmani (early Cretaceous) -- The first known definite monotreme, discovered in 1985.
Vincelestes neuquenianus (early Cretaceous, 135 Ma) -- A probably-placental mammal with some marsupial traits, known from some nice skulls. Placental-type braincase and coiled cochlea. Its intracranial arteries & veins ran in a composite monotreme/placental pattern derived from homologous extracranial vessels in the cynodonts. (Rougier et al., 1992)
Pariadens kirklandi (late Cretaceous, about 95 Ma) -- The first definite marsupial. Known only from teeth.
Kennalestes and Asioryctes (late Cretaceous, Mongolia) -- Small, slender animals; eyesocket open behind; simple ring to support eardrum; primitive placental-type brain with large olfactory bulbs; basic primitive tribosphenic tooth pattern. Canine now double rooted. Still just a trace of a non-dentary bone, the coronoid, on the otherwise all-dentary jaw. "Could have given rise to nearly all subsequent placentals." says Carroll (1988).
Cimolestes, Procerberus, Gypsonictops (very late Cretaceous) -- Primitive North American placentals with same basic tooth pattern.
So, by the late Cretaceous the three groups of modern mammals were in place: monotremes, marsupials, and placentals. Placentals appear to have arisen in East Asia and spread to the Americas by the end of the Cretaceous. In the latest Cretaceous, placentals and marsupials had started to diversify a bit, and after the dinosaurs died out, in the Paleocene, this diversification accelerated. For instance, in the mid- Paleocene the placental fossils include a very primitive primate-like animal (Purgatorius - known only from a tooth, though, and may actually be an early ungulate), a herbivore-like jaw with molars that have flatter tops for better grinding (Protungulatum, probably an early ungulate), and an insectivore (Paranyctoides).
The decision as to which was the first mammal is somewhat subjective. We are placing an inflexible classification system on a gradational series. What happened was that an intermediate group evolved from the 'true' reptiles, which gradually acquired mammalian characters until a point was reached where we have artificially drawn a line between reptiles and mammals. For instance, Pachygenulus and Kayentatherium are both far more mammal-like than reptile-like, but they are both called "reptiles".
1,931
posted on
02/11/2005 1:47:47 PM PST
by
shubi
(Peace through superior firepower.)
To: shubi
Actually, heat from the sun increases the entropy. As things are heated, molecules begin to move faster thereby increasing randomness. Ice, for example is much more ordered (less entropy) than water molecules in the air.
To: shubi
Shubi, you studied Hebrew for 13 years right?
To: Michael_Michaelangelo
I couldn't find any evidence, just assertion.
Point out one piece of scientific evidence against evolution on that site and we will discuss it in detail.
1,934
posted on
02/11/2005 1:51:56 PM PST
by
shubi
(Peace through superior firepower.)
To: Michael_Michaelangelo
are you asking to have your head handed to you on these?
I'm game, provided you agree to pay me for the effort.
1,935
posted on
02/11/2005 1:53:14 PM PST
by
King Prout
(Remember John Adam!)
To: Michael_Michaelangelo
That is an accusation not evidence against evolution.
Give us one piece of scientific evidence against evolution.
1,936
posted on
02/11/2005 1:53:54 PM PST
by
shubi
(Peace through superior firepower.)
To: Pantera
Science has plenty of evidence of speciation. So stop trying to wiggle out of the corner you have painted yourself into.
1,937
posted on
02/11/2005 1:55:09 PM PST
by
shubi
(Peace through superior firepower.)
To: Pantera
Nope, you don't understand the 2nd Law either.
The sun makes entropy irrelevant on Earth.
1,938
posted on
02/11/2005 1:56:36 PM PST
by
shubi
(Peace through superior firepower.)
To: Pantera
and, again, you dodge a reply from me to you, which answers one of your questions at least in part.
1,939
posted on
02/11/2005 1:57:02 PM PST
by
King Prout
(Remember John Adam!)
To: Tribune7
1,940
posted on
02/11/2005 1:57:08 PM PST
by
shubi
(Peace through superior firepower.)
Navigation: use the links below to view more comments.
first previous 1-20 ... 1,901-1,920, 1,921-1,940, 1,941-1,960 ... 2,241-2,242 next last
Disclaimer:
Opinions posted on Free Republic are those of the individual
posters and do not necessarily represent the opinion of Free Republic or its
management. All materials posted herein are protected by copyright law and the
exemption for fair use of copyrighted works.
FreeRepublic.com is powered by software copyright 2000-2008 John Robinson