
Posted on 02/20/2003 2:30:45 PM PST by Junior
IT STARTED with a biologist sitting on a grassy river bank in York, eating a sandwich. It ended in the discovery of a “scruffy little weed with no distinguishing features” that is the first new species to have been naturally created in Britain for more than 50 years.
The discovery of the York groundsel shows that species are created as well as made extinct, and that Charles Darwin was right and the Creationists are wrong. But the fragile existence of the species could soon be ended by the weedkillers of York City Council’s gardeners.
Richard Abbott, a plant evolutionary biologist from St Andrews University, has discovered “evolution in action” after noticing the lone, strange-looking and uncatalogued plant in wasteland next to the York railway station car park in 1979. He did not realise its significance and paid little attention. But in 1991 he returned to York, ate his sandwich and noticed that the plant had spread.
Yesterday, Dr Abbott published extensive research proving with DNA analysis that it is the first new species to have evolved naturally in Britain in the past 50 years.
“I’ve been a plant evolutionary biologist all my life, but you don’t think you’ll come across the origin of a new species in your lifetime. We’ve caught the species as it has originated — it is very satisfying,” he told the Times. “At a time in Earth’s history when animal and plant species are becoming extinct at an alarming rate, the discovery of the origin of a new plant species in Britain calls for a celebration.”
The creation of new species can takes thousands of years, making it too slow for science to detect. But the York groundsel is a natural hybrid between the common groundsel and the Oxford ragwort, which was introduced to Britain from Sicily 300 years ago. Hybrids are normally sterile, and cannot breed and die out.
But Dr Abbott’s research, published in the journal of the Botanical Society of the British Isles, shows that the York Groundsel is a genetic mutant that can breed, but not with any other species, including its parent species. It thus fits the scientific definition of a separate species.
“It is a very rare event — it is only known to have happened five times in the last hundred years” Dr Abbott said. It has happened twice before in the UK — the Spartina anglica was discovered in Southampton 100 years ago, and the Welsh groundsel, discovered in 1948.
The weed sets seed three months after germinating and has little yellow flowers. The species, which came into existance about 30 years ago, has been called Senecio eboracensis, after Eboracum, the Roman name for York. According to the research, it has now spread to spread to several sites around York, but only ever as a weed on disturbed ground.
However, more than 90 per cent of species that have lived subsequently become extinct, and its future is by no means certain.
“It is important for it to build up its numbers rapidly, or it could get rubbed out — which would be sad. The biggest threat to the new species is the weedkillers from the council,” Dr Abbott said.
However, he does not plan to start a planting programme to ensure his discovery lives on. “The next few years will be critical as to whether it becomes an established part of the British flora or a temporary curiosity. But we will let nature take its course,” he said.
You just don't get it, Mr. Smart-guy evil-u-sham-ist. All them big numbers was invented by a buncha evil people, probably athiests or commies, who hate God and want to destroy him. How else can you explain away the fact that Stegosaurus died for man's sins? Huh? Huh?
What would such a fossil look like? In other words, what is it that you would find convincing?
God invented creation // science !
Man invented lies // evolution !
The evidence you say does not exist actually does, although not in the quantity that you demand, because of the natural rarity of fossil-formation. But you didn't answer my question. What do you think a "transitional fossil" would look like?
The modern theory of evolution does not require gradual change. It in fact, the operation of Darwinian processes should yield exactly what we see in the fossil record. It is gradualism that we must reject, not Darwinism. ... Eldredge and I believe that speciation is responsible for almost all evolutionary change. Moreover, the way in which it occurs virtually guarantees that sudden appearance and stasis shall dominate the fossil record. All major theories of speciation maintain that splitting takes place rapidly in very small populations. The theory of geographic, or allopatric, speciation is preferred by most evolutionists for most situations (allopatric means "in another place"). A new species can arise when a small segment of the ancestral population is isolated at the periphery of the ancestral range. Large, stable central populations exert a strong homogenizing influence. New and favorable mutations are diluted by the sheer bulk of the population through which they must spread. They may build slowly in frequency, but changing environments usually cancel their selective value long before they reach fixation. Thus, phyletic transformation in large populations should be very rare — as the fossil record proclaims. But small, peripherally isolated groups are cut off from their parental stock. They live as tiny populations in geographic corners of the ancestral range. Selective pressures are usually intense because peripheries mark the edge of ecological tolerance for ancestral forms. Favorable variations spread quickly. Small peripheral isolates are a laboratory of evolutionary change.What should the fossil record include if most evolution occurs by speciation in peripheral isolates? Species should be static through their range because our fossils are the remains of large central populations. In any local area inhabited by ancestors, a descendant species should appear suddenly by migration from the peripheral region in which it evolved. In the peripheral region itself, we might find direct evidence of speciation, but such good fortune would be rare indeed because the event occurs so rapidly in such a small population. Thus, the fossil record is a faithful rendering of what evolutionary theory predicts, not a pitiful vestige of a once bountiful tale.
Stephen J. Gould, The Panda's Thumb: Reflections in Natural History, 1980, pp182-184
Yeah, the genes of the creature.
The part where you expanded on your answer and demonstrated how it supports your thesis seems to have gotten lost in transmission. Do try again.
Speaking of genes, though, they show *clear* evidence of common ancestry -- the kind you claim can't exist.
For example, genes can accidentally acquire and then retain chunks of virus DNA during viral infections. These imbedded bits of crud from past infections are then passed on to descendants, like any other genetic material in the chromosomal DNA. Since they can occur almost anywhere in the enormously long DNA strand (their placement during viral infection is random), two animals which have exactly the same viral "crud" fragment (out of a huge number of possible fragment sections and sizes) in exactly the same spot in the DNA strand is *extremely* strong evidence that they inherited that crud from a common ancestor, since independently acquired DNA garbage would be unlikely to happen in exactly the same spot in exactly the same way -- unlikely to the order of many billions-to-one odds.
Now let's look at a map of some common viral "crud" (known technically as "endogenous retroviruses") shared among various species:
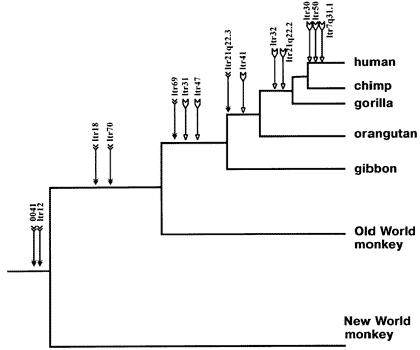
Oh, look -- 14 different endogenous retroviral sequences are found shared by various species in the primate family, *very strong* evidence of common ancestry. Even more interesting, the sequences which are shared (or not shared) by different species matches perfectly the "family tree" (i.e., which species split from which others in what order) indicated by the fossil record, morphological studies, and evolutionary predictions. Imagine that...
This sort of DNA support for evolution springs up EVERY time someone studies shared (or differing) DNA sequences among various species. Here's a similar map for various widely differing species of mammals:
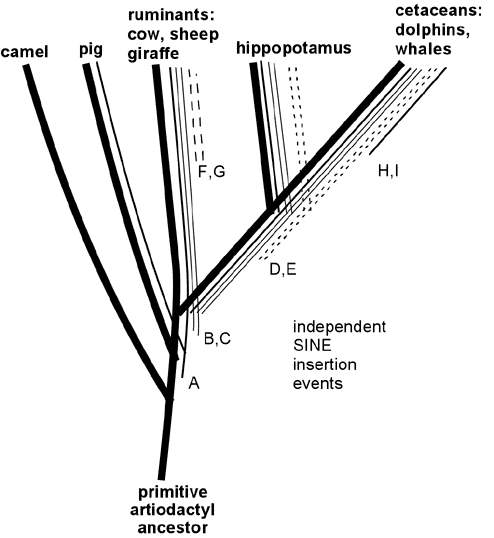
And here's a different study which discovered the same results:
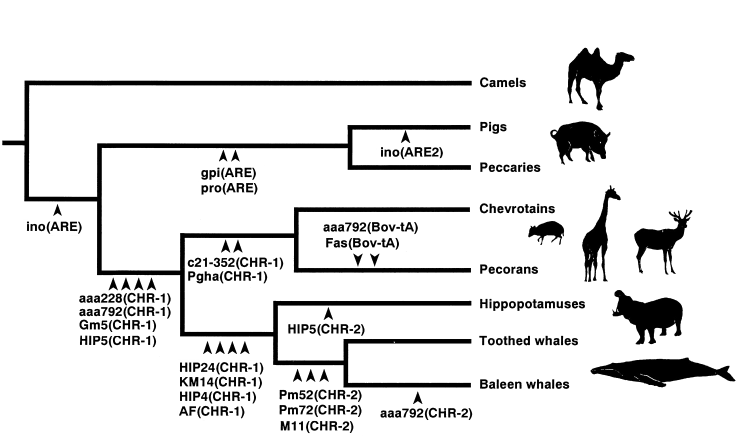
Here's the article for the above chart.
<< How does the fin know when to stop changing into a leg? Inquiring minds want to know. Evidently you do, so let's have it. >>
Fins don't evolve into legs.
They most certainly did. We have a pretty good stepwise sequence of fossils showing the transition from fins into legs:
Transition from primitive bony fish to amphibians
Few people realize that the fish-amphibian transition was not a transition from water to land. It was a transition from fins to feet that took place in the water. The very first amphibians seem to have developed legs and feet to scud around on the bottom in the water, as some modern fish do, not to walk on land (see Edwards, 1989). This aquatic-feet stage meant the fins didn't have to change very quickly, the weight-bearing limb musculature didn't have to be very well developed, and the axial musculature didn't have to change at all. Recently found fragmented fossils from the middle Upper Devonian, and new discoveries of late Upper Devonian feet (see below), support this idea of an "aquatic feet" stage. Eventually, of course, amphibians did move onto the land. This involved attaching the pelvis more firmly to the spine, and separating the shoulder from the skull. Lungs were not a problem, since lungs are an ancient fish trait and were present already.
- Paleoniscoids again (e.g. Cheirolepis) -- These ancient bony fish probably gave rise both to modern ray-finned fish (mentioned above), and also to the lobe-finned fish.
- Osteolepis (mid-Devonian) -- One of the earliest crossopterygian lobe-finned fishes, still sharing some characters with the lungfish (the other lobe-finned fishes). Had paired fins with a leg-like arrangement of major limb bones, capable of flexing at the "elbow", and had an early-amphibian-like skull and teeth.
- Eusthenopteron, Sterropterygion (mid-late Devonian) -- Early rhipidistian lobe-finned fish roughly intermediate between early crossopterygian fish and the earliest amphibians. Eusthenopteron is best known, from an unusually complete fossil first found in 1881. Skull very amphibian-like. Strong amphibian- like backbone. Fins very like early amphibian feet in the overall layout of the major bones, muscle attachments, and bone processes, with tetrapod-like tetrahedral humerus, and tetrapod-like elbow and knee joints. But there are no perceptible "toes", just a set of identical fin rays. Body & skull proportions rather fishlike.
- Panderichthys, Elpistostege (mid-late Devonian, about 370 Ma) -- These "panderichthyids" are very tetrapod-like lobe-finned fish. Unlike Eusthenopteron, these fish actually look like tetrapods in overall proportions (flattened bodies, dorsally placed orbits, frontal bones! in the skull, straight tails, etc.) and have remarkably foot-like fins.
- Fragmented limbs and teeth from the middle Late Devonian (about 370 Ma), possibly belonging to Obruchevichthys -- Discovered in 1991 in Scotland, these are the earliest known tetrapod remains. The humerus is mostly tetrapod-like but retains some fish features. The discoverer, Ahlberg (1991), said: "It [the humerus] is more tetrapod-like than any fish humerus, but lacks the characteristic early tetrapod 'L-shape'...this seems to be a primitive, fish-like character....although the tibia clearly belongs to a leg, the humerus differs enough from the early tetrapod pattern to make it uncertain whether the appendage carried digits or a fin. At first sight the combination of two such extremities in the same animal seems highly unlikely on functional grounds. If, however, tetrapod limbs evolved for aquatic rather than terrestrial locomotion, as recently suggested, such a morphology might be perfectly workable."
- Hynerpeton, Acanthostega, and Ichthyostega (late Devonian) -- A little later, the fin-to-foot transition was almost complete, and we have a set of early tetrapod fossils that clearly did have feet. The most complete are Ichthyostega, Acanthostega gunnari, and the newly described Hynerpeton bassetti (Daeschler et al., 1994). (There are also other genera known from more fragmentary fossils.) Hynerpeton is the earliest of these three genera (365 Ma), but is more advanced in some ways; the other two genera retained more fish- like characters longer than the Hynerpeton lineage did.
- Labyrinthodonts (eg Pholidogaster, Pteroplax) (late Dev./early Miss.) -- These larger amphibians still have some icthyostegid fish features, such as skull bone patterns, labyrinthine tooth dentine, presence & pattern of large palatal tusks, the fish skull hinge, pieces of gill structure between cheek & shoulder, and the vertebral structure. But they have lost several other fish features: the fin rays in the tail are gone, the vertebrae are stronger and interlocking, the nasal passage for air intake is well defined, etc.
More info on those first known Late Devonian amphibians: Acanthostega gunnari was very fish-like, and recently Coates & Clack (1991) found that it still had internal gills! They said: "Acanthostega seems to have retained fish-like internal gills and an open opercular chamber for use in aquatic respiration, implying that the earliest tetrapods were not fully terrestrial....Retention of fish-like internal gills by a Devonian tetrapod blurs the traditional distinction between tetrapods and fishes...this adds further support to the suggestion that unique tetrapod characters such as limbs with digits evolved first for use in water rather than for walking on land." Acanthostega also had a remarkably fish-like shoulder and forelimb. Ichthyostega was also very fishlike, retaining a fish-like finned tail, permanent lateral line system, and notochord. Neither of these two animals could have survived long on land.
Coates & Clack (1990) also recently found the first really well- preserved feet, from Acanthostega (front foot found) and Ichthyostega (hind foot found). (Hynerpeton's feet are unknown.) The feet were much more fin-like than anyone expected. It had been assumed that they had five toes on each foot, as do all modern tetrapods. This was a puzzle since the fins of lobe-finned fishes don't seem to be built on a five-toed plan. It turns out that Acanthostega's front foot had eight toes, and Ichthyostega's hind foot had seven toes, giving both feet the look of a short, stout flipper with many "toe rays" similar to fin rays. All you have to do to a lobe- fin to make it into a many-toed foot like this is curl it, wrapping the fin rays forward around the end of the limb. In fact, this is exactly how feet develop in larval amphibians, from a curled limb bud. (Also see Gould's essay on this subject, "Eight Little Piggies".) Said the discoverers (Coates & Clack, 1990): "The morphology of the limbs of Acanthostega and Ichthyostega suggest an aquatic mode of life, compatible with a recent assessment of the fish-tetrapod transition. The dorsoventrally compressed lower leg bones of Ichthyostega strongly resemble those of a cetacean [whale] pectoral flipper. A peculiar, poorly ossified mass lies anteriorly adjacent to the digits, and appears to be reinforcement for the leading edge of this paddle-like limb." Coates & Clack also found that Acanthostega's front foot couldn't bend forward at the elbow, and thus couldn't be brought into a weight-bearing position. In other words this "foot" still functioned as a horizontal fin. Ichthyostega's hind foot may have functioned this way too, though its front feet could take weight. Functionally, these two animals were not fully amphibian; they lived in an in-between fish/amphibian niche, with their feet still partly functioning as fins. Though they are probably not ancestral to later tetrapods, Acanthostega & Ichthyostega certainly show that the transition from fish to amphibian is feasible!
Hynerpeton, in contrast, probably did not have internal gills and already had a well-developed shoulder girdle; it could elevate and retract its forelimb strongly, and it had strong muscles that attached the shoulder to the rest of the body (Daeschler et al., 1994). Hynerpeton's discoverers think that since it had the strongest limbs earliest on, it may be the actual ancestor of all subsequent terrestrial tetrapods, while Acanthostega and Ichthyostega may have been a side branch that stayed happily in a mostly-aquatic niche.
In summary, the very first amphibians (presently known only from fragments) were probably almost totally aquatic, had both lungs and internal gills throughout life, and scudded around underwater with flipper-like, many-toed feet that didn't carry much weight. Different lineages of amphibians began to bend either the hind feet or front feet forward so that the feet carried weight. One line (Hynerpeton) bore weight on all four feet, developed strong limb girdles and muscles, and quickly became more terrestrial.
And if you're going to quibble about some of the details in the above, be sure to explain why no vertebrate had legs before the time of those fossils, then there arose amphibians with many fish-like features and primitive legs resembling the skeletal and muscular structure of the fish which existed shortly before... If the early amphibians didn't evolve from fish, God sure went out of his way to make it look as if they did.
You should have learned that when the coelecanth was rediscovered and what were thought to be transitional fin/legs turned out to still be 100% fins, supposedly 65 million years later.
What are you mumbling about here? No one ever claimed that the Coelacanth had legs. But fleshy lobed fins, like those in the Coelacanth, were an early stage on the way towards the development of legs.
Next you'll propose that sexual claspers on whales are vestigial legs.
Why would he want ot propose something false like that? Begone, straw man.
Which lack of transitional fossils were you referring to?
Was it the transition from fishes to amphibians?
Or was it the transition from amphibians to reptiles?
Or was it the transition from reptiles to birds?
Or was it the transition from reptiles to mammals?
Or was it the transition from a common ancestor to dogs, cats, bears, sealions, & hyenas?
Or was it the transition from land mammals to whales & dolphins?
Or was it the transition from a common ancestor to horses, donkeys, zebras, & rhinos?
Or was it the transition from a common ancestor to hippos, pigs, camels, & giraffes?
Or was it the transition from a common ancestor to modern humans & apes?
Sorry, but that's a lame argument. Technology is not evolutionary - which by definition must be the incidental (I started to say "accidental", but that's not quite right) acquisition of new characteristics due to necessity or convenience. Micro-evolution, whether you like the term or not, is the observable differentiation within a species, and is rarely seriously debated. Macro-evolution, on the other hand, is evolution across family or phylum lines.
Oh and btw, I'm not sure on the actual number but I'm quite sure the first computer had more vaccum tubes alone than your present day computer has parts.
Ummm... Not exactly. A vacuum tube is more analogous to a modern transistor than to a chip - which is composed of millions or billions of transistors (I lost track of the current record:-).
There are several dozens of fossils of our ancestors. You cite Nebraska Man (a very tentative claim that got blown up in the popular press before it was retracted). I'm sure at some point you'll mention Piltown Man (a hoax that was exposed as such by evolutionists) too.
But out here in the real world, the evidence shows several gradual transitions from Australopithecus to Homo sapiens sapiens. In fact some of them are so gradual that prominent creationist writers cannot agree among themselves over whether many fossils are "just an ape" or "just a human".
In fact, just today a new fossil from Olduvai Gorge is proving to yank Homo rudolfensis back into Homo habilis (the ancestor of Homo erectus).

Tonight at 6pm on RadioFR! Interviews with Grover Norquist, John Hager and Michael Zak! Plus, Doug from Upland interviews Ted Hayes, homeless advocate and strong supporter of military action in Iraq!
Click HERE to listen LIVE while you FReep! HIFI broadband feed HERE!
Why did you leave out speciation in your dividing line? You said microevolution is change within a species (which is true). This thread is all about the discovery of the evolution of a brand new species. Then you say macroevolution is the evolution of a new family.
Mainstream science calls change at the species level and above "macroevolution". Speciation is what has been observed here. That's macroevolution, because from now on the new species will only diverge further from its parent species.
The evos are pretty desperate, they'll take anything they can get - even a weed! But don't worry, now they will add hybrids to the theory of evolution. Who knows, sneezing may be the next proof of evolution!
No, they are not. But speciation is not enough to prove evolution. What evolution proposes is not speciation but TRANSFORMATION. What evolution needs to show is the creation of new features, functions, abilities, organs, systems, etc. No such proof has ever been found. A hybrid is no proof of evolution.
Disclaimer: Opinions posted on Free Republic are those of the individual posters and do not necessarily represent the opinion of Free Republic or its management. All materials posted herein are protected by copyright law and the exemption for fair use of copyrighted works.