4. Conclusions
The detailed evolutionary model described above is summarized in Figure 7. The role that various evolutionary processes played in the model can now be roughly quantified. Only one major shift of function occurred at the system level, the transition from a pilus to a protoflagellum. All of the other changes in system function can be seen as minor modifications of a basic function; if these are enumerated (export --> secretion --> adhesion --> pilus, and dispersal --> taxis), then four minor shifts of function occurred. In all cases a “shift” in function is actually more accurately described as an addition of function at the system level, as previous functions are maintained. At the level of subsystems (consisting of two or more proteins), the cooption events can be tabulated: subsystem cooption was invoked for the origin of the core export apparatus, outer membrane secretin (proto-FlgI) and lipoprotein chaperone (proto-FlgH), the adhesin ancestral to the axial protein family, the motor complex, and the chemotaxis/switch complex, for a total of five subsystem cooption events. In each of these cases, cooption occurred by the mutation of one protein to link two preexisting systems (Figure 7), followed by the duplication and integration of the new subsystem proteins into the major system. Except for the major transition between pilus and motility, subsystem cooption was associated with improvements of system function rather than major changes in system function. At the gene level, duplication events within the core system were invoked 11 times for origin of 12 axial proteins from one, and an additional time for the divergence of FliN and FliM. None of these events requires postulating functional shift at the subsystem or system levels. Addition of a new domain with novel functionality was identified twice (FliN+CheC --> FliM, rod cap+muramidase --> FlgJ), although it probably occurred in additional instances where homologies are currently more vague. It appears that loss of a component is only a possibility for the outer membrane secretin of the primitive type III secretion system, although if this became FlgI then no component loss events are necessary. This is the case even though some components that are ancient on the model (e.g., FliH) are apparently not absolutely required in modern flagella (Minamino et al., 2003). All other changes at all levels were matters of gradual improvement of function, i.e. optimization and co-adaptation of components. Even at this early stage of development, the model gives decent estimate of the relative importance of various evolutionary processes involved in the origin of complex biochemical systems.
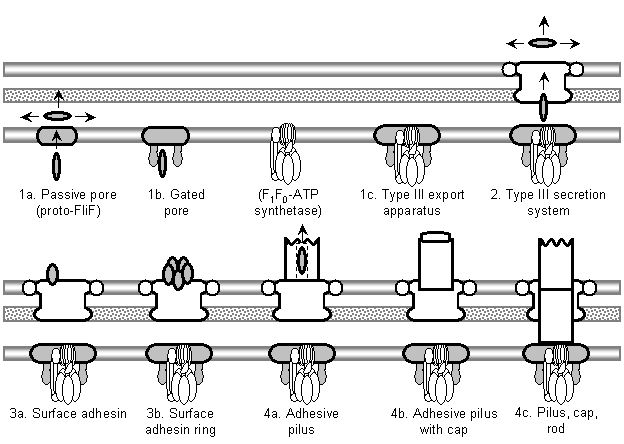
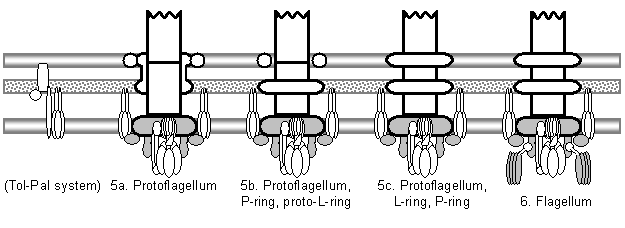
Figure 7: Summary of the evolutionary model for the origin of the flagellum, showing the six major stages and key intermediates. White components have identified or reasonably probable nonflagellar homologs; grey components have either suggested but unsupported homologs, or no specific identified homologs, although ancestral functions can be postulated. The model begins with a passive, somewhat general inner membrane pore (1a) that is converted to a more substrate-specific pore (1b) by binding of proto-FlhA and/or FlhB to FliF. Interaction of an F1F0-ATP synthetase with FlhA/B produces an active transporter, a primitive type III export apparatus (1c). Addition of a secretin which associates with the cytoplasmic ring converts this to a type III secretion system (2). A mutated secretion substrate becomes a secreted adhesin (or alternatively an adhesin is coopted by transposition of the secretion recognition sequence), and a later mutation lets it bind to the outer side of the secretin (3a). Oligomerization of the adhesin produces a pentameric ring, allowing more surface adhesins without blocking other secretion substrates (3b). Polymerization of this ring produces a tube, a primitive type III pilus (4a; in the diagram, a white axial structure is substituted for the individual pilin subunits; all further axial proteins are descended from this common pilin ancestor). Oligomerization of a pilin produces the cap, increasing assembly speed and efficiency (4b). A duplicate pilin that loses its outer domains becomes the proto-rod protein, extending down through the secretin and strengthening pilus attachment by association with the base (4c). Further duplications of the proto-rod, filament, and cap proteins, occurring before and after the origin of the flagellum (6) produce the rest of the axial proteins; these repeated subfunctionalization events are not shown here. The protoflagellum (5a) is produced by cooption of TolQR homologs from a Tol-Pal-like system; perhaps a portion of a TolA homolog bound to FliF to produce proto-FliG. In order to improve rotation, the secretin loses its binding sites to the axial filament, becoming the proto-P-ring, and the role of outer membrane pore is taken over by the secretin’s lipoprotein chaperone ring, which becomes the proto-L-ring (5b). Perfection of the L-ring and addition of the rod cap FlgJ muramidase domain (which removes the necessity of finding a natural gap in the cell wall) results in 5c. Finally, binding of a mutant proto-FliN (probably a CheC receptor) to FliG couples the signal transduction system to the protoflagellum, producing a chemotactic flagellum (6); fusion of proto-FliN and CheC produces FliM. Each stage would obviously be followed by gradual coevolutionary optimization of component interactions. The origin of the flagellum is thus reduced to a series of mutationally plausible steps.
Even the present extended treatment has left out detailed discussion of the origin of the chemotaxis and regulatory proteins listed in Table 2. However, many of these proteins have homologs functional in different systems, and the chaperones of axial proteins might have originated by duplication in a fashion similar to the axial proteins themselves. The evolution of the organization of flagellar genes and operons also deserves attention, although the precise organization found in modern bacteria is probably not essential (Kalir et al., 2001).